Article Text

Abstract
Objective In utero exposure of the fetus to Ro/La autoantibodies may lead to congenital heart block (CHB). In the mother, these autoantibodies are associated with activation of the type I interferon (IFN)-system. As maternal autoantibodies are transferred to the fetus during pregnancy, we investigated whether the type I IFN-system is activated also in newborns of anti-Ro/La positive mothers, and whether fetal IFN activation is affected by maternal immunomodulatory treatment.
Methods Blood drawn at birth from anti-Ro/La positive mothers, their newborns and healthy control pairs was separated into plasma and peripheral blood mononuclear cells (PBMC). PBMC were analysed directly or cultured. mRNA expression was analysed by microarrays, cell surface markers by flow cytometry, and IFNα levels by immunoassays.
Results We observed increased expression of IFN-regulated genes and elevated plasma IFNα levels not only in anti-Ro/La positive women, but also in their newborns. CD14+ monocytes of both anti-Ro/La positive mothers and their neonates showed increased expression of Sialic acid-binding Ig-like lectin-1, indicating cellular activation. Notably, the IFN score of neonates born to mothers receiving immunomodulatory treatment was similar to that of controls, despite persistent IFN activation in the mothers. In both maternal and neonatal PBMC, IFNα production was induced when cells were cultured with anti-Ro/La positive plasma.
Conclusions Ro/La autoantibody-exposed neonates at risk of CHB have signs of an activated immune system with an IFN signature. This study further demonstrates that neonatal cells can produce IFNα when exposed to autoantibody-containing plasma, and that maternal immunomodulatory treatment may diminish the expression of IFN-regulated genes in the fetus.
- autoantibodies
- autoimmune diseases
- Sjøgren's syndrome
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
Key messages
What is already known about this subject?
In utero exposure of the fetus to maternal Ro/La autoantibodies may lead to congenital heart block in the fetus, with recent studies indicating interferon (IFN) activation in fibroblasts of the fetal heart.
What does this study add?
This study demonstrates that Ro/La autoantibody-exposed neonates have signs of an activated immune system with an IFN signature and that maternal immunomodulatory treatment may diminish the IFN-induced gene expression in neonatal cells.
In this study we observe that neonatal cells can produce IFNα when exposed to autoantibody-containing plasma.
How might this impact on clinical practice?
Maternal treatment by immunomodulatory drugs has been suggested to protect against congenital heart block (CHB), and our study provides a molecular basis for such a potential protective effect.
Maternal IFN score in anti-Ro/La positive pregnancy could potentially be used as a biomarker for CHB risk.
Introduction
Systemic lupus erythematosus (SLE) and Sjögren’s syndrome (SS) are systemic autoimmune diseases characterised by multiorgan involvement and circulating autoantibodies against intracellular antigens such as Ro/SSA and La/SSB.1–3 These autoantibodies, in complex with ribonucleoproteins, can induce interferon (IFN)α production by plasmacytoid dendritic cells and peripheral blood mononuclear cells (PBMC).4 5 Accordingly, increased serum levels of type I IFN and overexpression of type I IFN-stimulated genes (IFN signature) are found in both SLE and SS.6 7 Type I IFN affects both the innate and adaptive immune systems and contributes to loss of tolerance and development of autoimmune diseases.8 9 Recently, a profound effect of type I IFN on central nervous system (CNS) neuronal synaptic mass was also reported.10 Commonly used therapies in both SLE and SS, such as hydroxychloroquine and glucocorticoids, have been suggested to diminish the type I IFN system activity, and several emerging drugs have been developed to directly target IFN-related pathways.8 9
During pregnancy, IgG is transported across the placenta, and Ro/SSA and La/SSB autoantibodies transfer to the fetus.11 12 The presence of these autoantibodies in the mother is associated with a risk of neonatal lupus erythematosus (NLE) in the child.13–15 The syndrome includes a congenital heart block (CHB), skin manifestations, liver and haematological involvement.16 17
It is well established that adult immune cells can produce type I IFN on exposure to Ro/SSA and La/SSB autoantibodies combined with apoptotic/necrotic material, but whether fetal cells are equally capable of such a response is not known. In addition, although the presence of Ro/SSA and La/SSB autoantibodies is known to associate with an IFN signature in SLE and SS patients, it has not been investigated if activation of the type I IFN system also occurs in fetuses exposed to Ro/SSA and La/SSB during pregnancy, and whether this may be affected by maternal immunomodulatory treatment. To address these questions, we sampled maternal and neonatal blood at birth from Ro/SSA and La/SSB autoantibody-positive, treated or untreated, pregnancies, as well as from control mother-neonate pairs, and analysed gene expression, cellular activation, and type I IFN levels in mothers and newborns.
Patients and methods
Study population
Pregnant anti-Ro/SSA and anti-La/SSB positive women under surveillance by Doppler echocardiography were recruited at the Department of Pediatric Cardiology, Karolinska University Hospital, Sweden (n=25). No mother gave birth to a baby with CHB. Healthy controls were recruited at the Labor and Delivery Unit at the same hospital (n=25). Blood was drawn from the mother and from the umbilical cord at delivery. Data on maternal diagnosis and medical treatment, as well as gestational age, sex, weight and length of the newborns are summarised in table 1. Samples available for respective analysis are defined in the figure legends.
Clinical characteristics of included mothers and neonates
Sample preparation
Blood samples were processed immediately after childbirth. Aliquots were first taken for flow cytometry and samples thereafter centrifuged at 1200 g for 10 min, plasma/serum collected, aliquoted and frozen at −80°C. PBMC were isolated with SepMate−50 tubes (Stemcell Technologies) according to the manufacturer’s instructions and frozen in fetal calf serum with 10% dimethylsulfoxide (DMSO) in liquid nitrogen. Aliquots of cells were also stored in RNA later solution (ThermoFisher Scientific) or Trizol (ThermoFisher Scientific).
RNA isolation
Cells stored in RNA later were diluted with 2× the volume of ice-cold PBS, cells stored in Trizol were extracted with chloroform. RNA was isolated using an RNAeasy mini kit according to the manufacturer’s instructions (Qiagen).
Microarrays and analysis
Analysis of RNA was performed at The BEA Core Facility, Department of Biosciences and Nutrition, Karolinska Institutet, using Affymetrix HTA V.2.0 microarrays. Differentially expressed genes were identified using the comparative marker selection module in GenePattern18 using a two tailed t-test with 0 permutations on log transformed data. Genes found to be differentially expressed between groups were visualised and hierarchically clustered using Morpheus (software.broadinstitute.org/morpheus/). The Molecular Signatures Database V.6.2 was used for Gene Set Enrichment Analysis (software.broadinstitute.org/gsea/msigdb) to find Gene Ontology (GO) terms over-represented among the top (p≤1E-04) differentially regulated genes and selected GO terms visualised using Morpheus. Pathway enrichment was visualised using the R package circlize. 19
IFN score
IFN scores were calculated as Z-scores as previously described,20 using 13 IFN-regulated genes (ISG15, IFI44, STAT1, PLSCR1, LY6E, IFIT1, IRF7, OAS1, OAS2, OASL, MX1, MX2 and IFIT3).
ELISA
Autoantibodies to Ro52, Ro60 and La were detected by ELISA (Orgenetec, Mainz, Germany) according to the manufacturer’s instructions. Samples were tested in duplicates at 1:500 and OD measured at 450 nm. Each plate contained at least two positive and two negative control sera.
IFNα assays
IFNα was measured by a dissociation-enhanced lanthanide fluoroimmunoassay (DELFIA) as previously described.21 In short, 96-well plates (Maxisorp, Fluoronunc) were coated with a one (supernatant analysis) or two (plasma analysis) of mouse anti-IFNα monoclonal antibodies (mAbs), LT27:273 (2 µg/mL) and LT27:293 (5 µg/mL), for capture, and a Europium-labelled anti-IFNα mAb LT27:297 (5 µg/mL) was used for detection. To reduce background, the dilution buffer (50 mM Tris-HCl, 0.9% NaCl, 0.5 mM CaCl2×2 H2O, 20 µM EDTA, 0.5% BSA, 0.005% NaN3, pH 7.8) was supplemented with an irrelevant murine mAb (50 µg/mL). After incubation with the mAbs, an enhancement solution (Perkin-Elmer) was added to the rinsed wells and fluorescence measured at 600 nm in a fluorometer (Wallac). The IFNα immunoassay standard was calibrated against the NIH reference leucocyte IFNα GA-23-902-530.
Flow cytometry staining and analysis
EDTA whole blood was incubated for 15 min at room temperature with green vidye dye (Thermofisher, ref. L23101) and an antibody cocktail containing: anti-sialic acid-binding Ig-like lectin 1 (SIGLEC-1) PE (clone 7–239, ref. 565248), anti-HLA-ABC PE-Cy5 (clone G46-2.6, ref. 555554), anti-CD3 PE-Cy7 (clone SK7, ref. 557851), anti-CD19 APC (clone HIB19, ref. 555415), anti-CD14 PB (clone M5E2, ref. 558121), anti-CD16 BV510 (clone 3G8, ref. 563829), all from Becton Dickinson, and anti-HLA-DR APC-Cy7 (clone L243, ref. 307618), BioLegend. Samples were incubated for 5 min at room temperature with RBC lysing buffer (Becton Dickinson, ref. 555899), the remaining leucocytes washed in PBS, and data acquired using a Gallios Flow Cytometer (Beckman Coulter). Gating strategy is visualised in online supplementary figure 1. Data analysis was performed with the FlowJo software (V.10.4.2, Tree Star).
Supplemental material
Cell cultures
PBMC (0.2×106) from neonates and their mothers were stimulated in 96-well flat-bottomed microtitre plates (Nunc, Denmark) with 0.1% (v/v) plasma, consisting of a pool of Ro/SSA and La/SSB positive plasma from three neonates and three mothers, respectively. The cell cultures were supplemented with 500 U/mL IntronA (Schering-Plough, Bloomfield, New Jersey, USA) and supernatant from necrotic cells prepared by freeze-thaw cycles as previously described,22 and incubated for 20 hours at 37°C with 5% CO2. IFNα in the supernatants was analysed by DELFIA immunoassay.21
Statistical analysis
Statistical analysis was performed using GraphPad PRISM V.6 software (San Diego, California, USA) except for microarray data. Differences between three groups were analysed with a one-way ANOVA and Tukey’s multiple comparison test, and two groups by an unpaired t-test when data passed a Kolmogorov-Smirnov test for normality. When normality could not be assumed, a Kruskal-Wallis test with Dunn‘s multiple comparisons test or Mann-Whitney test was used instead. Spearman’s rank correlation coefficient was calculated to assess correlation. The significance cut-off was set to p=0.05 throughout.
Results
Gene expression profile in neonates born to anti-Ro/La positive mothers
Gene expression analysis of PBMC purified from maternal blood at delivery showed significant differences in the expression of a large set of genes between the anti-Ro/La autoantibody-positive and control groups with 1795 genes passing a false discovery rate (FDR)≤0.1 (figure 1A). Notably, analysis of gene expression in neonatal cord blood-PBMC revealed that newborns of anti-Ro/La positive mothers also differentially expressed a high number of genes compared with newborns of healthy controls, with 1893 genes passing FDR≤0.1 (figure 1A). The overlap of differentially expressed genes in the maternal and neonatal groups was 910.
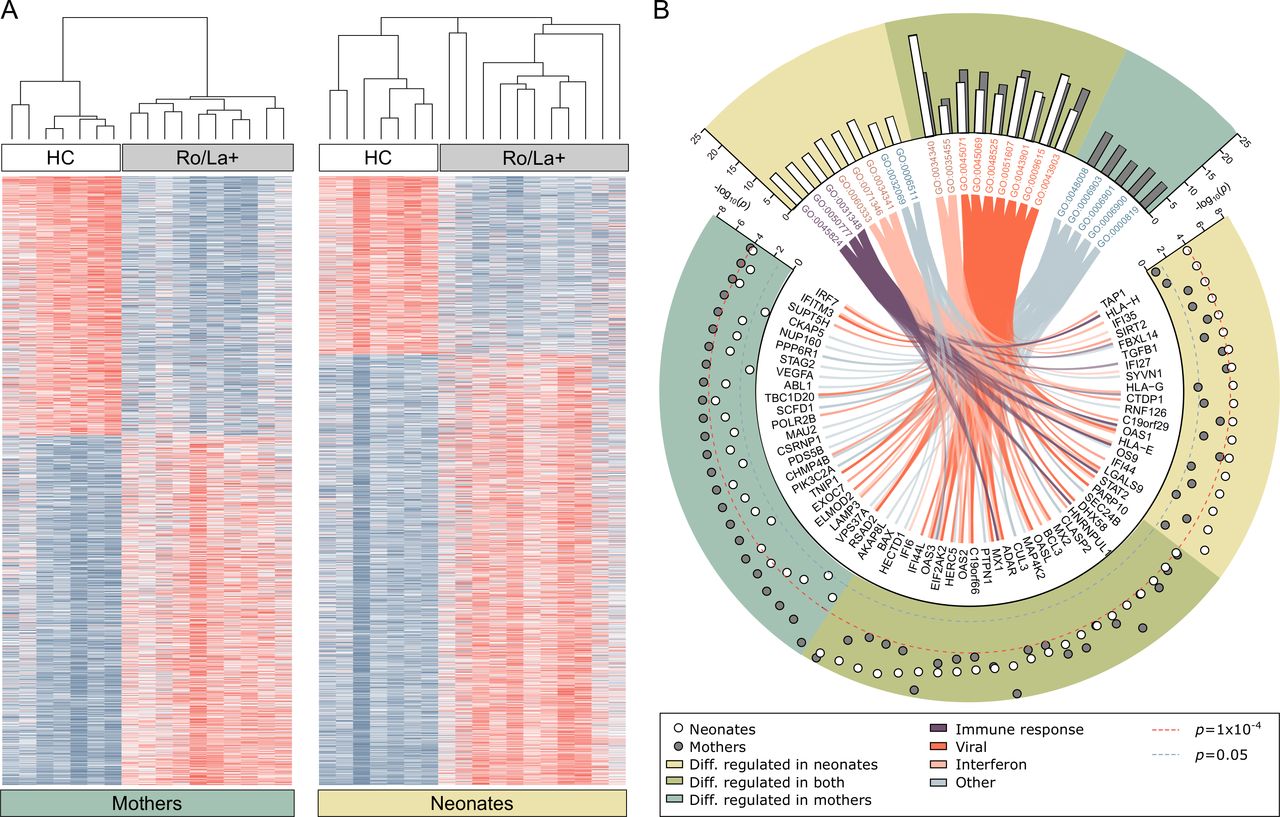
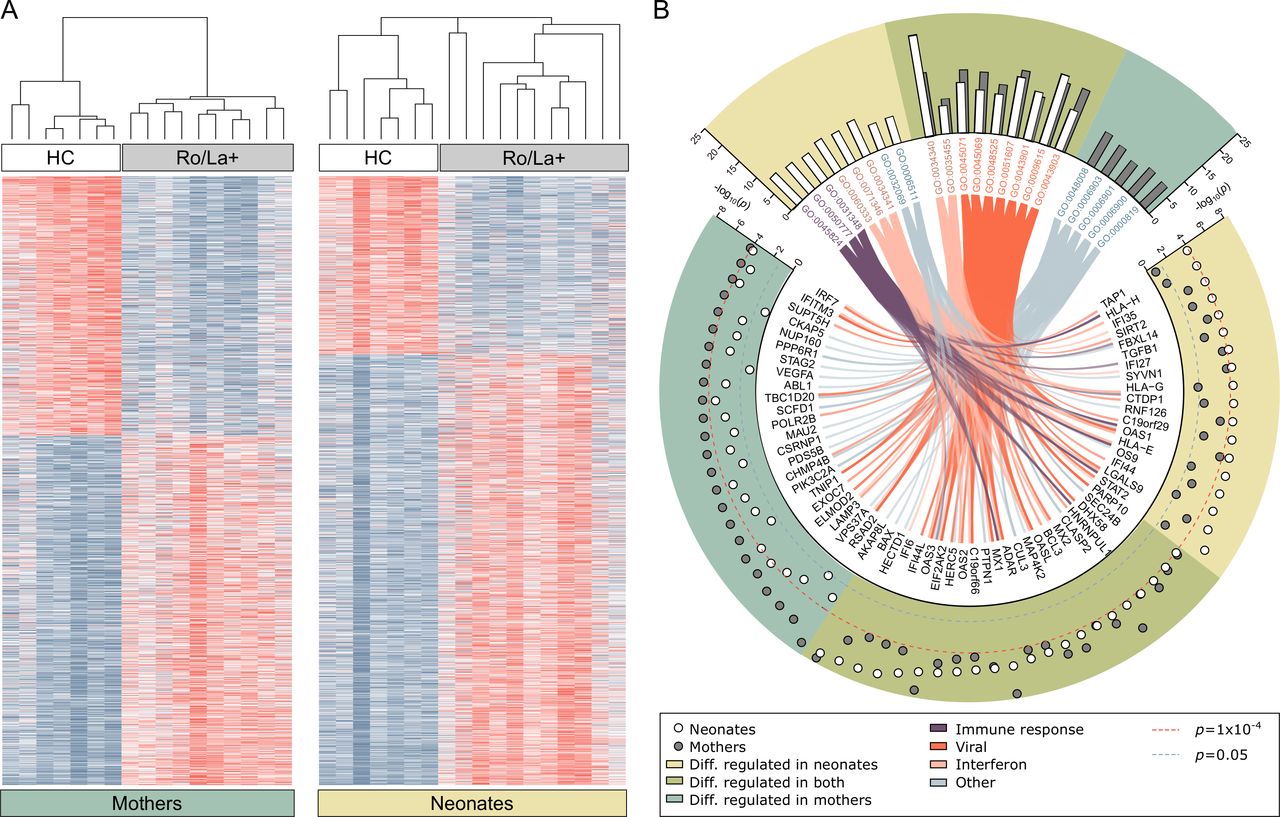
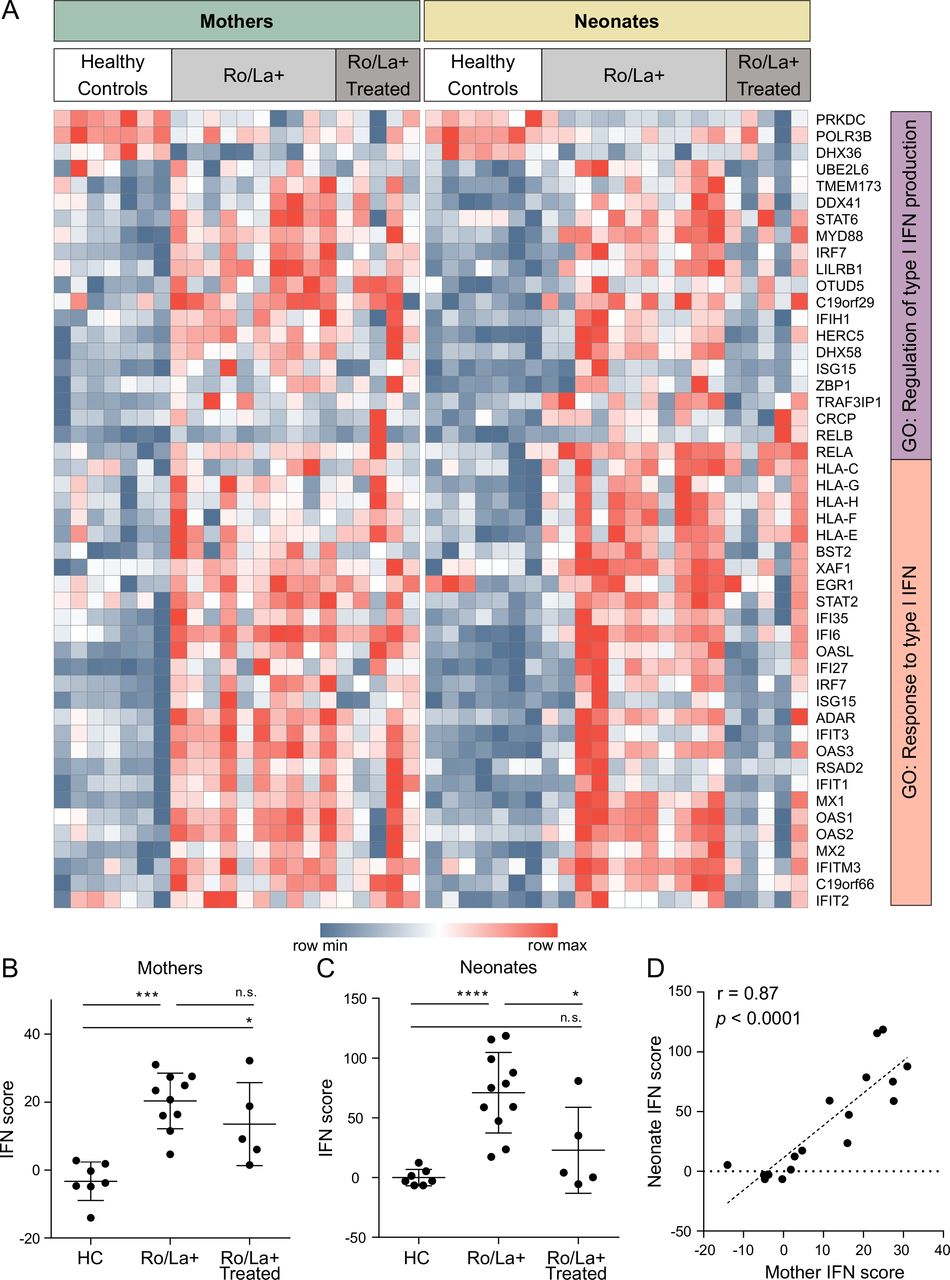
Gene expression differences in peripheral blood mononuclear cells of anti-Ro/La positive mothers, healthy control mothers and their respective neonates. (A) Heat map visualisation of genes differentially regulated between healthy control mothers and anti-Ro/La positive (Ro/La+) mothers without immunomodulatory treatment (FDR<0.1, n=1795) and in the newborns (neonates, cord blood samples) of control mothers versus newborns of Ro/La+mothers (FDR<0.1, n=1893). Genes and samples are visualised and hierarchically clustered using Morpheus. (B) The molecular signatures database was used for gene set enrichment analysis to find over-represented Gene Ontology (GO) terms representing the top differentially regulated genes (uncorrected p value cut-off of p<1E-04). The analysis identified 22 GO terms with more than 5% of the listed genes differentially regulated in either group, nine of those were over-represented in analysis of both mothers and newborns (green background), five only reached significance or the 5% cut-off in mothers (blue-green background) and eight only in the neonatal analysis (yellow background). The GO terms were manually grouped and colour coded and connected to their differentially regulated genes by the colour coded chords. White circles represent the transformed, uncorrected p value from the comparative marker selection comparison of Ro/La+ and healthy control neonates, grey circles represent the same for the mothers. Red dotted line marks the inclusion cut-off for the enrichment analysis, blue line represents the uncorrected significance cut-off of p<0.05. Genes significantly differentially regulated in either analysis belonging to any of the pictured GO terms are depicted. RNA of sufficient amount and quality was available for anti-Ro/La+mothers #1–10, newborns #1–11 and seven healthy control mother-newborn pairs (table 1). HC, healthy control.
To understand which pathways the identified genes represent, the top differentially regulated genes (p≤0.0001), including 238 genes from the maternal analysis and 197 genes from the neonatal analysis, were used for gene set enrichment analysis looking for over-representation of GO terms (figure 1B). The analysis revealed significant enrichment in GO terms predominantly relating to IFN signalling and responses, including GO terms representing viral responses, in both mothers and neonates (figure 1B, online supplementary tables S1 and S2).
Supplemental material
These results demonstrate for the first time that PBMC derived from anti-Ro/La-exposed neonates have a significantly different gene expression profile compared with neonates born to healthy controls, with differentially expressed genes predominantly involved in IFN pathways, similar to what is observed in their anti-Ro/La positive mothers.
Neonates born to untreated anti-Ro/La positive mothers, but not neonates born to women receiving immunomodulatory drugs, display a high IFN score at birth
Specific analysis of IFN-regulated genes revealed the presence of an IFN signature in anti-Ro/La positive mothers, consistent with previous reports of an IFN signature in patients with SLE and SS (figure 2A). Importantly, the expression of IFN-induced genes in PBMC isolated from the cord blood of anti-Ro/La-exposed infants was also increased compared with controls (figure 2A). We next used the expression data of IFN-induced genes to calculate an IFN score.20 We observed that anti-Ro/La positive women had a significantly higher IFN score than healthy controls (figure 2B), and, importantly, that their neonates also had a significantly higher IFN score compared with control neonates (figure 2C), with maternal and neonatal IFN scores showing a strong and significant positive correlation (figure 2D).

Interferon (IFN) regulated genes differentially expressed in peripheral blood mononuclear cells of anti-Ro/La positive mothers, healthy control mothers and their newborns. (A) Expression of genes present in the go terms ‘Gene Ontology (GO): regulation of type I IFN production’ and ‘GO: response to type I IFN’ was filtered for differential expression (p<0.05) in the comparative marker selection for healthy controls versus Ro/La positive (Ro/La+) mothers and newborns. Genes and samples were visualised using Morpheus. (B) IFN scores of the mothers (healthy controls, Ro/La+ without or with immunomodulatory treatment. (C) IFN scores of the neonates (born by healthy control, Ro/La+ without or with immunomodulatory treatment mothers). (D) IFN scores plotted for mothers (non-treated only) versus their neonates, showing a significant correlation between IFN scores in mothers and their neonates at birth (r=0.87, p<0.0001). The figure includes data from Ro/La+mothers #1–10, 12–16, newborns #1–16, and seven healthy control mother-newborn pairs (table 1). ****p<0.0001, ***p<0.001, **p<0.01, *p<0.05.
Subsequent analysis of anti-Ro/La positive mothers treated with immunomodulatory drugs and their neonates revealed that, while the mothers’ IFN scores were not different from those of the untreated group of mothers, the expression of IFN-induced genes in their neonates was significantly lower compared with neonates of untreated mothers (figure 2B,C). Notably, there was no significant difference in the IFN score of neonates born to anti-Ro/La positive mothers treated with immunomodulatory drugs and neonates born to healthy controls (figure 2C).
Altogether, our data reveal that not only mothers with circulating anti-Ro/La autoantibodies, but also their newborns, have a significantly higher IFN score than controls, and that maternal immunomodulatory treatment may reduce the expression of IFN-regulated genes in the fetus.
Neonates born to anti-Ro/La positive women have elevated plasma IFNα levels and increased monocyte SIGLEC-1 expression at birth
To assess if the observed increased expression of IFN-induced genes in PBMC of neonates born to anti-Ro/La positive mothers was associated with the presence of type I IFN in their plasma at birth, a DELFIA assay17 that detects most IFNα subtypes was performed on cord blood. This revealed significantly higher plasma levels of IFNα in babies born to anti-Ro/La positive mothers compared with controls, paralleling what was observed in the mothers (figure 3A).
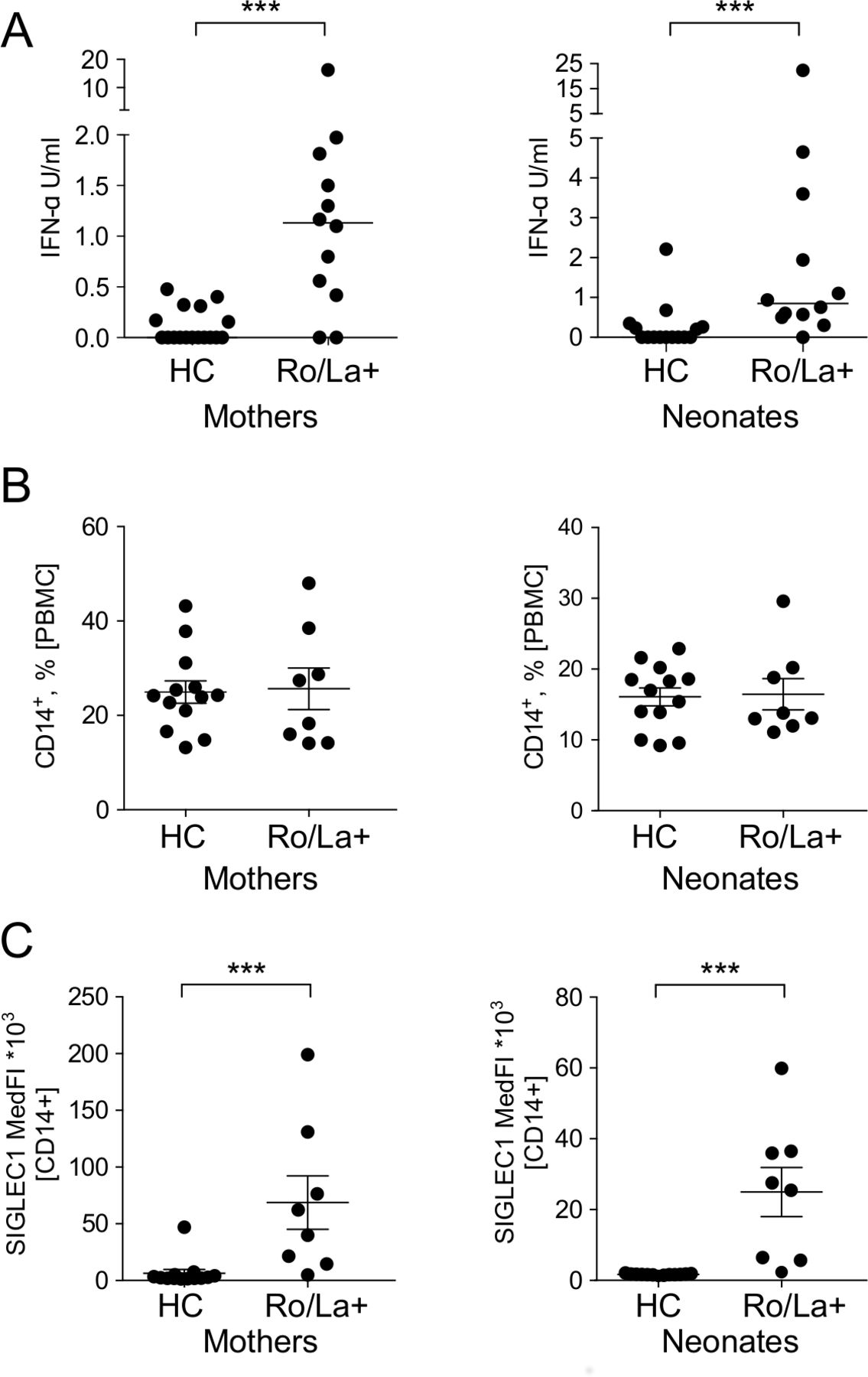
Interferon (IFN)α levels and immune activation in newborns exposed to Ro/La autoantibodies in utero. (A) IFNα levels in the plasma of mothers and their newborns (Ro/La+mothers #1–11, 25, newborns #1–11, 25 and 17 HC mothers, 15 HC newborns, table 1). (B,C) Frequency of CD14+ monocytes and SIGLEC-1 expression (median fluorescence intensity; MedFI) in the mothers and their newborns (Ro/La+mothers #17–24, newborns #17–24 and 13 healthy control mother-newborn pairs, table 1). ***p<0.001. HC, healthy controls; PBMC, peripheral blood mononuclear cells; SIGLEC-1, sialic acid-binding Ig-like lectin 1.
In CHB, macrophages have been identified as a major cell type in the cardiac lesions,23 and CD68+ macrophages have also been observed in NLE skin lesions.24 Expression of SIGLEC-1, a monocyte activation marker, is induced by type I IFN, and maternal SIGLEC-1 expression on monocytes has recently been described as a risk factor for the development of autoimmune CHB.25 SIGLEC-1-positive macrophages have also recently been suggested to play a role in the CHB inflammatory process in the fetal heart via activation of type I IFN.25 26 We therefore analysed SIGLEC-1 expression on circulating CD14+ monocytes from cord blood in anti-Ro/La positive and control pregnancies. While the frequency of CD14+ monocytes did not differ between the two groups (figure 3B), expression of SIGLEC-1 was significantly increased in neonates born to anti-Ro/La positive mothers compared with controls, suggesting activation of monocytes in neonates at risk of NLE (figure 3C).
Collectively, our data show that the presence of an IFN signature in neonates born to anti-Ro/La positive mothers is accompanied by elevated IFNα levels and signs of activated monocytes in their circulation, and support a potentially functionally important role for the type I IFN system in NLE pathogenesis.
IFNα production is readily induced in neonatal cells in response to neonatal plasma from anti-Ro/La positive pregnancies
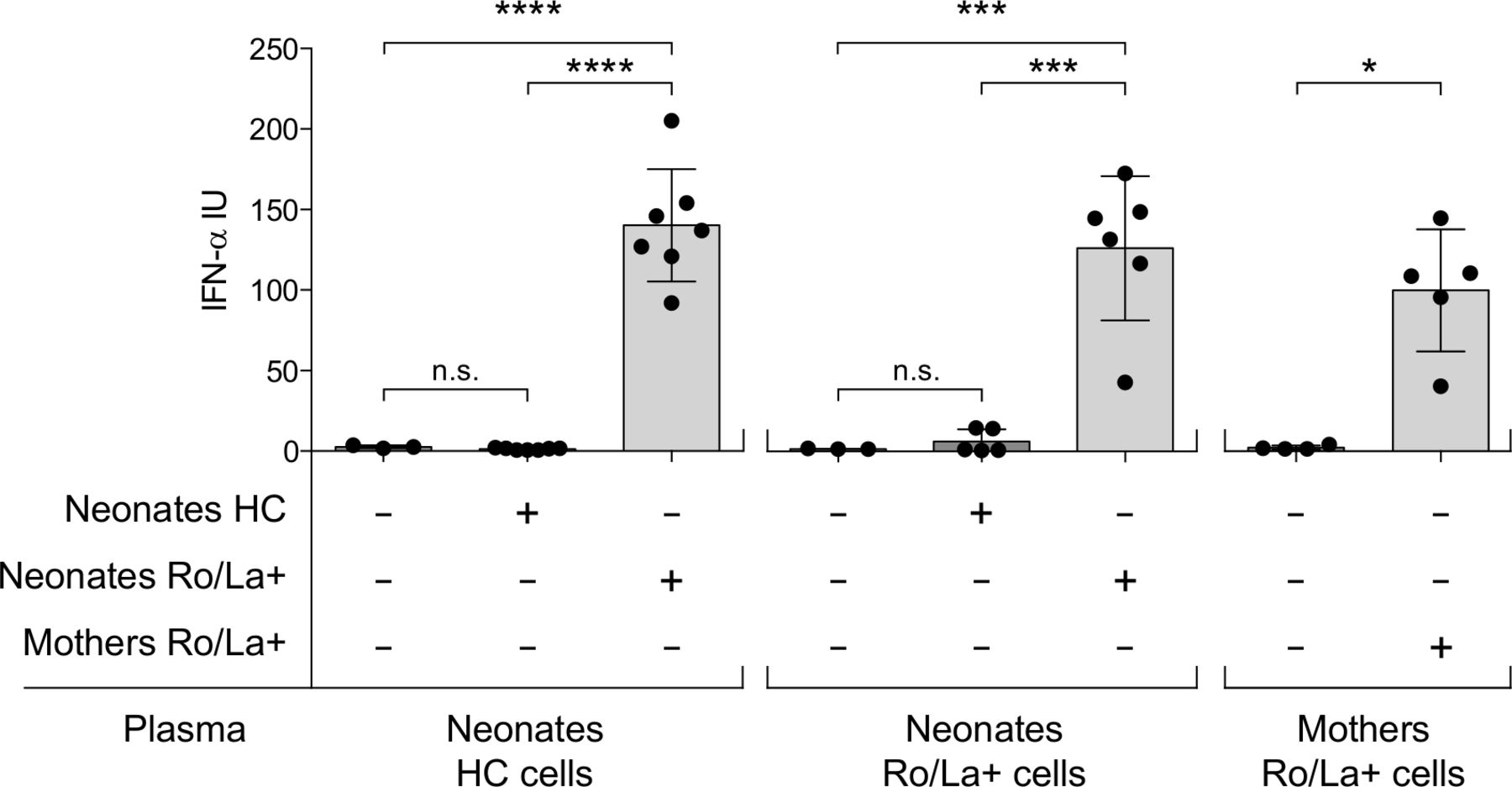
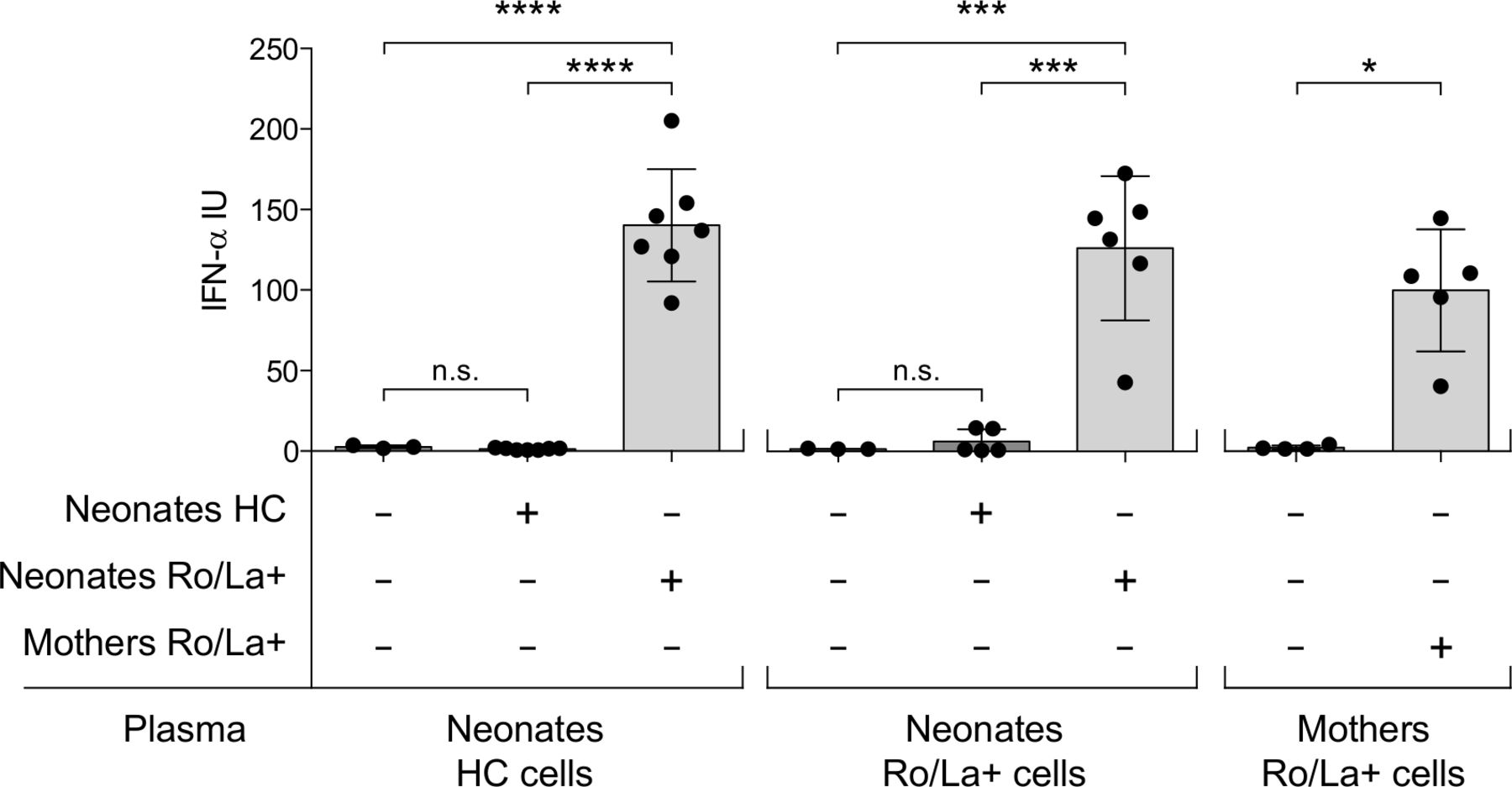
Adult immune cells can produce type I IFN after exposure to Ro/La antibodies combined with apoptotic/necrotic material, but whether fetal immune cells also have that ability is unknown. To understand whether the circulating IFNα we observed in neonates born to anti-Ro/La positive mothers could have been produced by the fetal immune cells, we cultured PBMC from cord blood with plasma combined with necrotic material and measured the production of IFNα. We primarily used PBMC isolated from cord blood of neonates born by healthy controls as we hypothesised that there is no previous IFN-activation in the fetus when Ro/La-autoantibody exposure occurs, but investigated also cells from in utero anti-Ro/La-exposed newborns as well as maternal cells for comparison. Cells were exposed to plasma from newborns of anti-Ro/La-positive mothers, plasma of newborn controls, or anti-Ro/La positive maternal plasma.
No cells produced IFNα in response to plasma from control newborns. PBMC from both control and anti-Ro/La-exposed newborns however readily produced high levels of IFNα in response to plasma from anti-Ro/La-exposed neonates. The IFNα levels produced were similar to those produced by maternal PBMC exposed to maternal anti-Ro/La positive plasma (figure 4).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Interferon (IFN)α production is readily induced in neonatal peripheral blood mononuclear cells (PBMC) in response to plasma from anti-Ro/La positive pregnancies. IFNα produced by PBMC isolated from HC and anti-Ro/La autoantibody exposed neonates, as well as mothers, and cultured under indicated differential conditions. ****p<0.0001, ***p<0.001, *p<0.05. HC, healthy control.
In all, these data demonstrate for the first time that neonatal PBMC can produce IFNα following exposure to Ro/La antibody containing plasma.
Discussion
In this study, we explored the activation of the type I IFN system in Ro/SSA and La/SSB autoantibody-positive pregnancies, and found that not only the mothers but also their newborns have elevated plasma IFNα levels, a cellular IFN-signature and activated monocytes. We further demonstrate that fetal PBMC can produce IFNα in response to Ro/La antibody-positive plasma, and, notably, that maternal immunomodulatory treatment may diminish the IFN-response in the fetus.
The circulating IFNα we detected in the anti-Ro/La-exposed newborns may derive from either the mother or be produced by fetal cells. In this report, we show for the first time that fetal immune cells are capable of producing IFNα in response to Ro/La autoantibody-positive plasma. It is therefore possible that maternal Ro/La autoantibodies, previously shown to induce IFNα production in adult PBMC and demonstrated to cross the placenta,11 12 induce IFNα production by fetal cells in vivo. The possible sources for the fetal IFNα are not mutually exclusive, but in vitro studies have shown that proinflammatory cytokines, including IFNα, do not pass normal term placenta.27 28 The placenta of early and/or autoimmune pregnancies may still functionally differ,29 potentially allowing the transfer of maternal IFN. Our observation that newborns of mothers on immunomodulatory treatment have significantly lower IFN scores than newborns of untreated mothers, although the IFN score in treated mothers were still high, however, supports the hypothesis that at least a major part of the IFN is produced in the fetus.
The role of IFNα in the pathogenesis of NLE, including CHB, is only beginning to be understood. Interestingly, recent studies have indicated that cardiac tissue injury in the fetus may be mediated by an effect of type I IFN on fetal fibroblasts.30 These studies proposed a role for SIGLEC-1-positive leucocytes, detected in fetal hearts, in mediating cardiac tissue inflammation and fibrosis.26 Another recent publication demonstrated that mothers of children with CHB have a significantly higher expression of SIGLEC-1 on monocytes and elevated levels of circulating IFNα,25 in line with our findings. In this report, we additionally show that increased expression of SIGLEC-1 on CD14+ monocytes is not confined to mothers but is also observed in their newborns. It has previously been reported that being exposed to Ro/SSA and La/SSB as well as other autoantibodies during pregnancy influences neurodevelopment, both in children with and without CHB.31 Specific anti-NMDA autoantibodies have been linked to CNS lupus,32 but a recent report demonstrating IFN-dependent synapse loss and behavioural phenotypes in lupus-prone mice suggests that type I IFN per se may influence neurodevelopment.10 This emphasises the importance of further studies to understand the possible effect of type I IFN in NLE, and on the development and outcome of the fetus.
Maternal hydroxychloroquine treatment has been suggested to protect against CHB development.33 34 Our observation of an IFN signature in neonates born to untreated anti-Ro/La positive mothers but not in neonates of mothers treated with immunomodulatory drugs may provide a molecular basis for such a potential protective effect. Hydroxychloroquine crosses the placenta and has been shown to inhibit endosomal toll-like receptor (TLR) signalling, and may thus decrease an autoantibody-dependent type I IFN activation in the fetus. Interestingly, our data indicate that the immunomodulatory treatment could have an impact on the IFN score of the fetus even if that of the mother remains high, which suggests that hydroxychloroquine and other immunomodulatory treatment may benefit the fetus even if the maternal IFN score is not brought back to normal levels. Notably, several studies indicate that a substantial number of children with CHB are born to anti-Ro/La positive mothers without a rheumatic diagnosis and consequently no treatment.17 35–37
The study is limited by not including Ro/La autoantibody-exposed neonates with CHB. However, our main aim was to assess the potential presence of an IFN signature in Ro/La autoantibody-exposed infants, and the long-standing high-dose steroid treatment commonly administered in pregnancies complicated by CHB will affect cytokine production, masking fetal CHB-related gene expression. Activation of type I IFN pathways has however been demonstrated in cardiac-derived cells from a few hearts of fetuses with CHB in terminated pregnancies where treatment was not initiated,26 30 suggesting that type I IFN is indeed relevant to the pathogenesis of CHB. Prospective studies characterising maternal IFN levels and IFN scores in early stages of anti-Ro/La positive pregnancies will be of interest to understand their potential predictive value for CHB development.
In summary, we here demonstrate that neonates born to anti-Ro/La positive mothers present elevated IFNα levels and an IFN signature in peripheral blood at birth, and that type I IFN can be produced by neonatal immune cells after exposure to Ro/La autoantibody containing plasma. Our findings uncover a new molecular pathway through which anti-Ro/La autoantibodies may contribute to the risk of NLE and CHB. In addition, our data suggest that maternal immunomodulatory treatment might prevent activation of the type I IFN system in the fetus.
Acknowledgments
We thank Dr Birgitta Gullstrand for excellent technical assistance and the Maternity wards at the Karolinska University Hospital, The South Hospital, the Danderyd University Hospital, BB Sophia, the Skåne University Hospital, the Örebro University Hospital, the Sahlgrenska University Hospital, the University Hospital Uppsala for invaluable support in blood sampling. We gratefully acknowledge Dr Aurélie Ambrosi, Karolinska Institutet, for contributing to the writing of the paper.
References
Footnotes
MH and GET are joint first authors.
Contributors MH, GET and MW-H conceived the study. MH, GET, MI, VO, NK, LL, JT, AS, AAB, EA, KG, LR, GB, AE, S-ES, M-LE performed experiments, contributed to data acquisition and analysis. MH, GET and MW-H took the lead in writing the manuscript. All authors provided critical review and approval of the final version manuscript.
Funding This study was supported by grants from the Swedish Research Council, the Heart-Lung Foundation, the Stockholm County Council, the Karolinska Institute, the Swedish Rheumatism Association, King Gustaf the V:th 80-year Foundation, the Swedish Society of Medicine, the Ingegerd Johansson donation, the Foundation Samariten and the Torsten and Ragnar Söderberg Foundation.
Competing interests None declared.
Patient consent for publication The mothers gave written informed consent.
Ethics approval The study was approved by the Local Ethics Committee Stockholm.
Provenance and peer review Not commissioned; externally peer reviewed.
Data availability statement All data relevant to the study are included in the article or uploaded as supplementary information.