Article Text

Abstract
Objective To identify the role of mature nerve growth factor (mNGF), its immature form proNGF and their receptors in arthritis inflammation.
Methods Real-time PCR, western blot and ELISA were performed to evaluate NGF, proNGF, their receptor and cytokine expression in synovial tissue and cells of patients with juvenile idiopathic arthritis (JIA) and rheumatoid arthritis (RA), and controls.
Results proNGF and not mNGF is the prevalent form measured in synovial fluids of patients with JIA and RA with synovial fibroblasts as a major source of proNGF in the inflamed synoviae. p75NTR, the specific receptor for proNGF, is the NGF receptor most expressed in mononuclear cells of patients with JIA, while TrkA is the prevalent receptor in healthy donors. In ex vivo experiments the effects of proNGF differ from those of mNGF, suggesting that the balance of p75NTR and TrkA expression represents a critical factor in regulating mNGF/proNGF functions, determining which intracellular pathways and biological activities are triggered. Contrary to NGF, proNGF administration increased inflammatory cytokines but not interleukin (IL)-10 expression, inducing a stronger activation of p38 and JNK pathways. proNGF effects depend on its binding to p75NTR, as inhibition of p75NTR with neutralising antibodies or LM11A-31 abolished proNGF-induced production of IL-6 in patients’ mononuclear cells, while inhibition of TrkA did not. There is a correlation in patients with arthritis between high p75NTR levels and severity of clinical symptoms.
Conclusions Our data suggest that an active proNGF-p75NTR axis promotes proinflammatory mechanisms contributing to chronic tissue inflammation, and that the use of p75NTR inhibitors may represent a new therapeutic approach in chronic arthritis.
- inflammation
- juvenile idiopathic arthritis
- rheumatoid arthritis
- cytokines
- synovial fluid
This is an Open Access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/
Statistics from Altmetric.com
Key messages
What is already known about this subject?
High levels of nerve growth factor (NGF) have been demonstrated in patients with arthritis, but there is no clear evidence of NGF-specific effects on the inflammatory response.
Moreover, no information is available on the role of the other NGF form, the proNGF, and its preferential receptor system p75NTR/sortilin in inflammatory diseases.
What does this study add?
This study shows for the first time that in inflamed synoviae the high levels of proNGF, with activated synovial fibroblasts as its major source, and high levels of p75NTR, the specific receptor of proNGF, correlate with the severity of clinical symptoms in patients with arthritis.
How might this impact on clinical practice?
The in vitro data showing that binding of proNGF to p75NTR enhances inflammatory cytokine production and that inhibition of p75NTR activity reduces inflammation suggest that inhibitors of p75NTR could represent a new therapeutic approach in chronic arthritis.
Introduction
High levels of nerve growth factor (NGF) characterise inflamed tissues of patients with a variety of chronic inflammatory diseases,1–3 but how NGF and the activation of its receptors, TrkA and p75NTR, regulate cells and mediators during inflammatory responses is not yet well defined.
We recently demonstrated in human monocytes that inflammatory stimuli, while activating a proinflammatory response through Toll-like receptors (TLR), also upregulate the expression of the NGF receptor TrkA.4 TrkA activation inhibits TLR signalling, and this results in a decrease in inflammatory cytokines and a concomitant increase in anti-inflammatory cytokine production. A marked downregulation of TrkA expression characterises mononuclear cells of patients with juvenile idiopathic arthritis (JIA), who showed an altered NGF regulatory feedback mechanism: in contrast to observations in healthy donors, the addition of NGF did not reduce interleukin (IL)-6 production from JIA mononuclear cells.4
To assess whether there were other anomalies in the NGF pathways of patients with JIA, in this study we investigated the modulation of the other NGF receptor, p75NTR, during the inflammatory response in patients with arthritis and how its engagement affected inflammation. Initially defined as the low-affinity NGF receptor, p75NTR was believed to bind to TrkA, increasing both TrkA specificity and affinity for NGF, thus contributing to the creation of a high-affinity NGF receptor.5 Currently, the role of p75NTR appears to be much more complex, and studies on its effects and signalling pathways, restricted mainly to cells of the nervous system, have not fully defined it. This is quite surprising, considering that p75NTR was the first characterised member of the tumour necrosis factor (TNF)-receptor superfamily,6 whose members share structural similarities, such as an intracellular ‘death domain’ and the lack of intrinsic enzymatic activity, and play a crucial role in the homeostasis of the immune system.7 8 As in other members of this receptor family, p75NTR has been reported to have different, or even opposite, effects depending on environment and cell type. Indeed, studies on neuronal cells have shown that p75NTR can either induce apoptosis or favour survival in differentiating neurons and affects cytoskeleton organisation, neurite growth and axonal degeneration.9 This variety of effects is mediated by many cytosolic interactors and a growing number of coreceptors.9 10 Recently, much attention has been paid to sortilin, a transmembrane receptor belonging to the Vps10p domain-receptor family11 that, in addition to its role in regulating protein trafficking, specifically binds proNGF, the immature form of NGF, increasing the specificity and affinity of p75NTR for proNGF.12
Studies on neurons have suggested that proNGF is not an inactive precursor,13 but acts as a signalling ligand with effects that are distinct from or even opposite to those of mature nerve growth factor (mNGF).14 Notably, in vivo studies have shown that proNGF can be even more abundant than mNGF in brain and in peripheral tissues.15 16
No information is available on the effects of the binding of proNGF to p75NTR in regulating the immune response. The main aim of this study was to investigate the potential involvement of the proNGF-p75NTR axis in chronic arthritides and the effects of proNGF through the p75NTR receptor on mononuclear cells from inflamed joints. Our results indicate that mononuclear cells of patients with JIA show an altered p75NTR/TrkA balance that, together with the high concentration of proNGF found in inflamed synovia, contributes to activate inflammatory pathways, leading to enhanced production of inflammatory cytokines. The blocking of p75NTR in ex vivo experiments with JIA mononuclear cells strongly suggests that the relative levels of p75NTR expression play a key role in the inflammatory response and provide the rationale for specifically targeting p75NTR for the treatment of chronic inflammatory diseases.
Materials and methods
Patients
Diagnosis of JIA and classification into JIA subtypes was based on the International League of Associations for Rheumatology classification criteria. Patients were classified as persistent oligoarticular, extended oligoarticular or polyarticular JIA. The median age at disease onset was 3.0 years (IQR 2.1–6.7) and the median disease duration at sampling was 5.0 years (IQR 1.9–8.7). All patients had active disease at the time of sampling. Patients receiving systemic glucocorticoids were not included in the study. Approximately 50% of the patients were receiving methotrexate with or without associated TNF inhibitors, with the remaining patients being treated with non-steroidal anti-inflammatory drugs only. Incidentally, no differences in p75NTR expression were found among the two groups. Synovial fluid samples were obtained at the time of intra-articular steroid injection. The study was approved by the Ethical Committee of the Bambino Gesù Children’s Hospital and written consent was obtained from parents of children as appropriate. Peripheral blood from healthy children of comparable age who underwent routine blood sampling prior to minor elective surgery for malformations were used as controls. Synovial tissue was obtained from patients with rheumatoid arthritis (RA) undergoing ultrasound-guided synovial biopsy at the Division of Rheumatology of the IRCCS Policlinico S Matteo Foundation, Pavia, Italy. Fibroblast-like synoviocytes (FLS) were obtained from patients with RA and osteoarthritis (OA) after synovial biopsy.17 Fibroblasts obtained from skin of healthy donors were used as controls.
Cell cultures
Mononuclear cells were isolated from peripheral blood (PBMC) and from synovial fluids (SFMC) of patients with JIA by Ficoll-Hypaque (Pharmacia, Uppsala, Sweden). Cells were cultured in AIM V Serum Free Medium (Life Technologies, Rockville, Maryland, USA) either without the addition of any stimuli (unstimulated, US) or activated with 3 ng/mL Lipopolysaccharide (LPS, Sigma Aldrich, St Louis, Missouri, USA). Mononuclear cells were treated with or without mNGF or mutated cleavage-resistant proNGF (Alomone Labs, Jerusalem, Israel) at a concentration, respectively, of 100 ng/mL and 200 ng/mL, calculated to yield the same molar concentration as described in neurons.18 For this study we selected an endotoxin-free proNGF to avoid unspecific activation of mononuclear cells. To block p75NTR and TrkA activity, the cells were incubated with either 2.5 µg/mL affinity-purified anti-p75NTR antibody (Millipore, Billerica, Massachusetts, USA) or 10 nM LM11A-31 (Dompé Farmaceutici, L’Aquila, Italy) or 3 µg/mL affinity-purified anti-TrkA antibody (R&D Systems, Minneapolis, Minnesota, USA) for 1 hour and then stimulated with 2 ng/mL LPS with or without the addition of 200 ng/mL proNGF. FLS was obtained from synovial tissue of patients with RA (RA FLS) or patients with OA (OA FLS) after enzymatic digestion with 0.2% collagenase type IV in high-glucose Dulbecco's Modified Eagle Medium (DMEM) supplemented with 10% fetal calf serum and antibiotics. Isolated synovial cells were expanded in DMEM supplemented with 10% FCS and used between the third and sixth passage. All fibroblasts were then cultured for 18 hours at a concentration of 50 000 cells/mL in serum-free Medium 106 (Thermo Fisher Scientific, Waltham, Massachusetts, USA) only.
RNA extraction and real-time PCR analysis
Total RNA, extracted from cells and tissues using TRIzol Reagent (Life Technologies), was used for first-strand cDNA synthesis (SuperScript VILO cDNA Synthesis Kit; Invitrogen, Carlsbad, California, USA). Freshly purified mononuclear cells obtained from peripheral blood or synovial fluid after Ficoll gradient centrifugation were used to measure mRNA expression levels of TrkA, p75NTR and sortilin in patients with JIA and healthy donors (controls CTRL) (figure 1A,B) and to associate them with disease activity (figure 3). FLS obtained from RA FLS, from OA FLS or control fibroblasts obtained from skin of healthy donors (CTRL FB) were used to analyse NGF mRNA expression levels and were untreated (figure 2D). For cytokine mRNA expression (figure 4C), mononuclear cells from synovial fluid (SFMC) or from peripheral blood (PBMC) of patients with JIA and mononuclear cells from peripheral blood of healthy controls were cultured for 3 hours before RNA extraction in AIM V serum-free medium only (US conditions) or treated with LPS (3 ng/mL), with or without the addition of mutated proNGF (200 ng/mL) or mNGF (100 ng/mL). Real-time PCR was performed on the ABI PRISM 7900 HT Sequence Detector (Applied Biosystems, Foster City, California, USA) platform, using TaqMan Universal Master Mix (Applied Biosystems). TrkA, p75NTR, NGF, sortilin, IL-1β, IL-6, IL-8, IL-10 and TNF-α mRNA expressions were tested using Assays on Demand reagents (TrkA Hs01021011_m1; p75NTR Hs00182120_m1; NGF Hs00171458_m1; sortilin Hs00361760_m1; IL-1β Hs00174097_m1; IL-6 Hs00985639_m1; IL-8 Hs00174103_m1; IL-10 Hs00961622_m1; TNF-α Hs99999043_m1; Applied Biosystems). TaqMan Endogenous Control human GAPDH (Hs99999905_m1; Applied Biosystems) was used as housekeeping gene. Relative quantification was performed using the comparative Ct method and results were expressed in arbitrary units (AU). Expression levels were calculated as 2−ΔCt and then compared with each other, while fold changes were calculated using the 2−ΔΔCt equation.19

p75NTR, TrkA and sortilin expression levels in patients with JIA. (A) mRNA expression levels of TrkA, p75NTR and sortilin in freshly isolated mononuclear cells from peripheral blood of healthy children (CTRL PBMC) and in mononuclear cells from peripheral blood or from synovial fluids of patients with JIA (respectively, JIA PBMC and JIA SFMC) were quantified by real-time PCR analysis. JIA PBMC (n=34) and JIA SFMC (n=66) express low TrkA and high p75NTR mRNA levels compared with CTRL PBMC (n=10). Sortilin, a coreceptor for p75NTR essential for proNGF binding, is highly expressed in JIA SFMC (n=19) and expressed in JIA PBMC (n=13) and in CTRL PBMC (n=7). The results are expressed as arbitrary units (AU) after normalisation with the housekeeping gene GAPDH. (B) p75NTR mRNA levels are higher in SFMC than in PBMC of matched patients with JIA (n=18). (C) Sortilin mRNA levels are higher in SFMC than in PBMC of matched patients with JIA (n=11). (D) Western blot and densitometric analysis of three independent experiments confirmed that p75NTR was the prevalent NGF receptor in JIA mononuclear cells , while TrkA was the most expressed NGF receptor in CTRL PBMC. Sortilin is also highly expressed in JIA mononuclear cells. *p<0.05, **p<0.01, ***p<0.001. JIA, juvenile idiopathic arthritis; NGF, nerve growth factor; PBMC, mononuclear cells isolated from peripheral blood; SFMC, mononuclear cells isolated from synovial fluids.

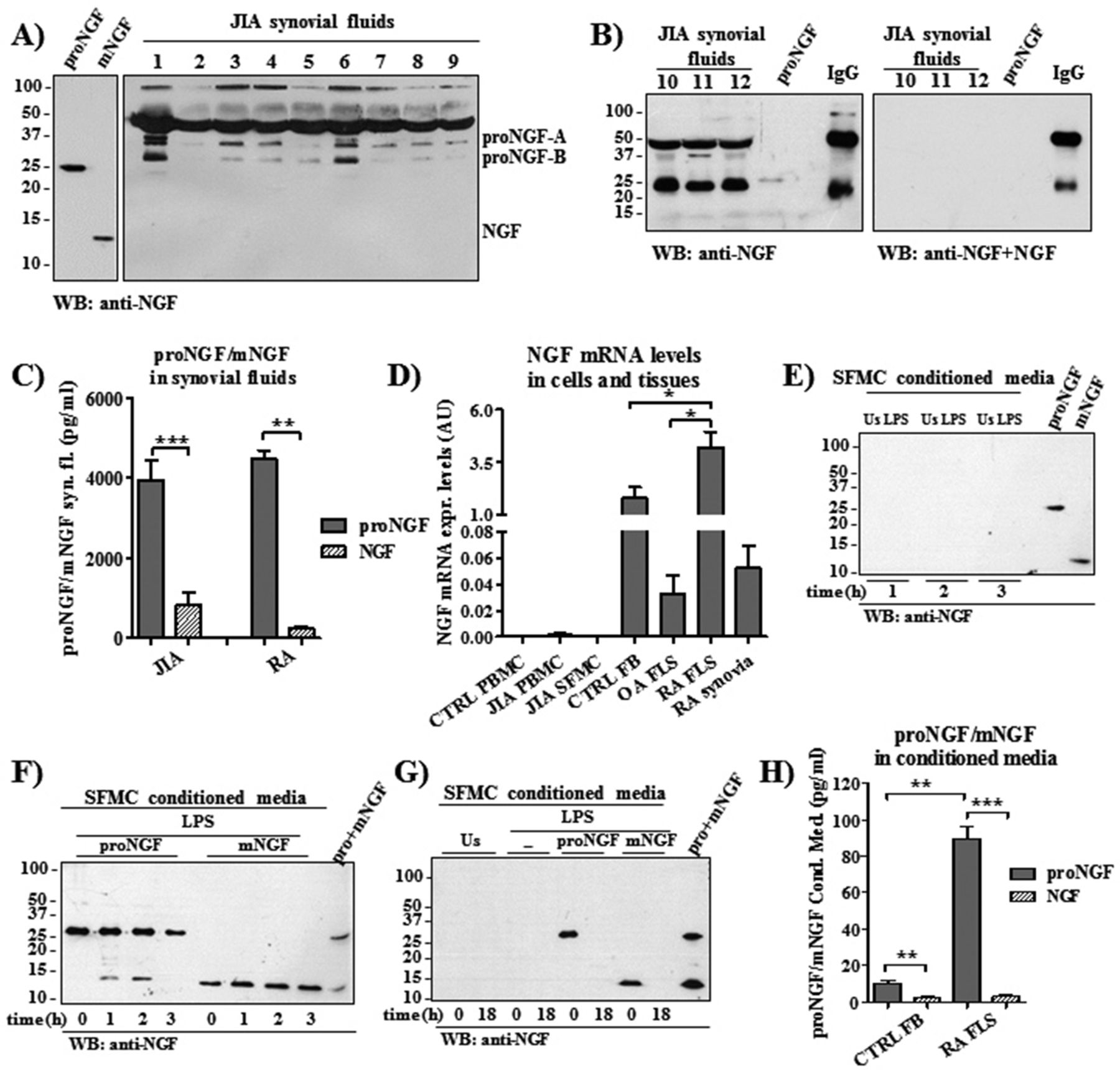
Mature and immature NGF forms in patients with JIA and RA. (A,B) Only proNGF forms of different molecular weights but not mature NGF were detected by western blot in synovial fluids of 12 patients with JIA (40 μg total protein). (B) Blocking the anti-NGF antibody by adding mature NGF results in the disappearance of the specific proNGF bands observed in synovial fluids of three patients with JIA, as well as of the band of the 25 kDa commercial proNGF that was added as positive control. (C) In JIA and RA synovial fluids, proNGF and mature NGF (mNGF) concentrations were assessed using a newly developed ELISA showing that proNGF is 4.8-fold higher in JIA (n=27) and 16.8-fold higher in patients with RA (n=5) than mNGF. (D) Real-time PCR shows a very low expression of NGF mRNA in mononuclear cells obtained from peripheral blood of healthy donors (CTRL PBMC) or in mononuclear cells from peripheral blood (JIA PBMC) and from synovial fluids of patients with JIA (JIA SFMC). High NGF mRNA expression levels characterised synovial tissues (RA synovia; n=5). Fibroblast-like synoviocytes (FLS; n=3) of patients with RA express more NGF mRNA than FLS from osteoarthritis patients (OA FLS) (n=4) and control fibroblasts (CTRL FB; n=4). Results were expressed as arbitrary units (AU) and obtained after normalisation with the housekeeping gene glyceraldehyde 3-phosphate dehydrogenase (GAPDH). (E) Both unstimulated (US) and LPS-stimulated JIA SFMC do not release proNGF or mNGF in the supernatants. Commercial proNGF and mNGF were added as positive controls. (F) Western blot shows the stability of exogenous added mNGF and proNGF. After its addition to culture media, proNGF is still detected after 3 hours and undergoes a minor maturation into mNGF in the first 2 hours of incubation (left side of F). mNGF is not degraded in the first 3 hours after its addition (right side of F). Commercial proNGF and mNGF were included as positive controls. (G) Neither proNGF nor mNGF were released in culture media of US or LPS-stimulated JIA SFMC after 18 hours of incubation. Exogenous proNGF or mNGF were detected only at time of supplementation (in agreement with F) but were no longer detectable after 18 hours of incubation. Commercial proNGF and mNGF were included as positive controls. (H) ELISA instead shows that conditioned media of RA FLS (n=5) have higher concentrations of proNGF than control fibroblasts (CTRL FB; n=5) and that the concentration of proNGF is higher than mNGF after 18 hours of incubation. *p<0.05, **p<0.01, ***p<0.001. JIA, juvenile idiopathic arthritis; LPS, Lipopolysaccharide; NGF, nerve growth factor; PBMC, mononuclear cells isolated from peripheral blood; RA rheumatoid arthritis; SFMC, mononuclear cells isolated from synovial fluids.
Cytokine and mNGF-proNGF ELISA
Conditioned media were collected from mononuclear cells obtained from peripheral blood (PBMC) and from synovial fluid (SFMC) of patients with JIA and from RA, OA and skin fibroblasts (FLS) after 18 hours of incubation in synthetic medium without any kind of stimulation (US). Conditioned media of mononuclear cells of patients with JIA and healthy donors treated with LPS, with or without the addition of either proNGF or mNGF, were also collected and all conditioned media were stored at −80°C. Human IL-6 was measured using ELISA kit (R&D Systems). Concentrations of mNGF and proNGF were analysed using specific ELISAs developed by the Consiglio Nazionale delle Ricerche (CNR) group and recently described.20 In brief, to recognise mNGF and proNGF the AF-556 capture antibody (R&D Systems) was coated overnight at room temperature (RT). After blocking for 1 hour at RT with phosphate buffered saline (PBS) +1% Bovine Serum Albumin (BSA), samples were incubated for 2 hours at RT. The microwells were then incubated with EP1318Y-proNGF (Chemicon) or 27/21-mNGF (Chemicon) detection monoclonal antibodies dissolved in blocking buffer for 2 hours at RT. Horseradish peroxidase (HRP)-conjugated antibodies (Cell Signaling, Danvers, Massachusetts, USA) were added and incubated for 1 hour at RT. To visualise antibody reactivity, the chromogenic substrate 3’,3’,5,5’-tetramethylbenzidine (TMB, Sigma Aldrich) was used and colour development was stopped by adding 1N HCl. The colourimetric reaction was measured in absorbance mode at 450 nm by a Multiskan EX ELISA reader (Thermo Fisher Scientific Laboratory Equipment, Hudson, New Hampshire, USA).
Western blot analysis
Mononuclear cells from synovial fluid (SFMC) of patients with JIA in AIM V serum-free medium were either treated with 3 ng/mL LPS and 200 ng/mL proNGF for 5 or 10 min or not treated (US). For p75NTR inhibition, cells were starved for 3 hours in Roswell Park Memorial Institute medium (RPMI) 0.1% FCS, preincubated with 10 nM LM11A-31 for 1 hour in AIM V serum-free medium and then stimulated with LPS, with or without the addition of 50 ng/mL NGF or 100 ng/mL mutated-proNGF for 5 min. After cell lysis in ice-cold RIPA buffer (Cell Signaling), equal amounts of protein were resolved by Sodium Dodecyl Sulphate - PolyAcrylamide Gel Electrophoresis (SDS-PAGE) and electrotransferred to nitrocellulose membranes. The total protein loading for synovial fluids (see figure 2A,B) was assessed by Ponceau staining (not shown) and gel analysis by the National Institutes of Health (NIH) ImageJ software. The amount of loaded proteins for synovial fluids was found to be almost equal for all the samples run on SDS-PAGE (coefficient of variation=5.81%). Nitrocellulose membranes (Amersham Life Sciences, Little Chalfont, UK) were probed with antibodies against phospho-stress-activated protein kinase/c-Jun NH(2)-terminal kinase (SAPK/JNK) (Thr183/Tyr185), and total SAPK/JNK, phospho-p38 MAPK (Thr180/Tyr182) and total p38 MAPK (all from Cell Signaling), p75NTR (clone 8211) (EMD Millipore, Billerica, Massachusetts, USA), TrkA (sc-118) and GAPDH (all from Santa Cruz Biotechnology, Dallas, Texas, USA) and tubulin (Sigma Aldrich). To evaluate the relative concentrations of proNGF and mNGF in synovial fluids of patients with arthritis, we used the rabbit polyclonal antibody mNGF H-20 (SantaCruz sc-548 WB) that recognises immunoreactive NGF forms ranging from 13 to 150 kDa after SDS-PAGE in reducing conditions.20 Nitrocellulose-bound antibodies were detected by chemiluminescence with ECL (Amersham Life Sciences).
Statistical analysis
Statistical analysis of the results was performed using non-parametric tests (GraphPad Prism 5 Software, La Jolla, California, USA). Data are presented as mean±SEM or SD as appropriate, and the level of statistical significance was set at p<0.05.
Results
p75NTR, sortilin and proNGF expression in patients with JIA and RA
In patients with JIA (n=70) the p75NTR receptor is highly expressed: the mean expression of p75NTR was twofold higher in mononuclear cells from peripheral blood (PBMC) and 19-fold higher in mononuclear cells from synovial fluids (SFMC) than in control PBMC (p<0.05 and p<0.001, respectively, Figure 1A). The analysis of paired samples of PBMC and SFMC in 18 patients with JIA showed that SFMCs are characterised by a significant upregulation of p75NTR expression (p<0.001) when compared with PBMC of the same patients (Figure 1B). Confirming our previous results on a smaller group of patients,4 the expression of TrkA in PBMC from patients with JIA was significantly lower than in control PBMC (p<0.01) and, even at the site of inflammation, TrkA expression levels in SFMC remained significantly lower (p<0.05) than those of healthy children (Figure 1A). The concomitant increase in p75NTR expression and decrease in TrkA expression determined an abnormal receptor balance: in PBMC of healthy donors the p75NTR/TrkA expression ratio was 0.5±0.09, while in patients with JIA it was 2.4±0.11 in PBMC (p<0.01) and 13.1±1.1 in SFMC (p<0.001). Consistently, western blot clearly indicated that p75NTR, and not TrkA, was the predominant receptor in JIA mononuclear cells, in contrast to control PBMC, where TrkA, and not p75NTR, is the prevalent NGF receptor (Figure 1D). Moreover, in agreement with real-time PCR data, JIA SFMCs express higher p75NTR protein levels than JIA PBMCs (Figure 1D). Mononuclear cells also express sortilin, the p75NTR coreceptor, and its expression level was markedly increased in JIA SFMCs, which also have the highest p75NTR expression (Figure 1A). Similarly to p75NTR, in paired JIA samples sortilin expression is significantly higher (p<0.01) in SFMC than in PBMC (Figure 1C). In western blot, JIA mononuclear cells showed higher levels of sortilin than healthy controls (Figure 1D).
Since studies on neuronal cells have shown that sortilin and p75NTR coexpression increases the specificity and affinity of p75NTR to proNGF,12 contributing to the formation of high-affinity receptors for NGF, we investigated whether the preferential ligand of p75NTR, that is proNGF, was present in the synovial fluids of patients with JIA and RA. Using an anti-NGF antibody that recognises both mature and immature human NGF forms,20 we found that the predominant form of NGF in the synovial fluid of patients with JIA was proNGF, while mNGF was not detectable by western blot (Figure 2A,B). In agreement with previous reports,20–22 we found that the majority of proNGF forms detectable in synovial fluid weighed 50, 34 and 25 kDa, the two latter probably representing the proNGF-A and proNGF-B splicing variants, respectively,21 23 while the 50 kDa could represent a glycosylated proNGF24 or a dimer of the proNGF-B. To quantify more accurately the concentration of both proNGF and mNGF, we used ELISAs that specifically recognise either the immature (ie, proNGF) or the mature form of human NGF.20 Confirming the western blot results, the concentration of proNGF far exceeded that of mNGF in synovial fluids of patients with JIA and RA (Figure 2C).

p75NTR and disease severity (A) in persistent oligoarticular (n=29), extended oligoarticular (n=16) and polyarticular (n=13) patients p75NTR mRNA expression levels are higher in patients with JIA with the highest number of inflamed joints (**p<0.01). (B) p75NTR mRNA levels found in SFMC of patients with JIA (n=57) correlate with the number of active joints (rs=0.43; p=0.001) and (C) with C reactive protein levels (n=50; rs=0.39; p=0.006). JIA, juvenile idiopathic arthritis; SFMC, mononuclear cells isolated from synovial fluids.
In order to identify the cellular source of proNGF, we found that NGF mRNA expression was very low in PBMC and SFMC from patients, while in PBMC from healthy donors (n=8) NGF expression was not detectable in our real-time PCR experimental conditions (Figure 2D). The protein analysis showed that SFMC did not release proNGF or mNGF in the culture media, neither in unstimulated conditions nor after LPS stimulation (Figure 2E). In the conditioned media of LPS-treated cells, the only detectable NGF forms were those exogenously added (Figure 2F,G). In order to assess the stability of mNGF and of the cleavage-resistant proNGF in our culture conditions, we collected conditioned media at different time points (Figure 2F,G). We found that up to 3 hours after LPS activation (Figure 2F), the time at which we collected cells for real-time PCR analysis of cytokine expression, there is any appreciable degradation of proNGF and mNGF, while neither NGF form was detectable after 18 hours of incubation (Figure 2G). This suggests that proNGF and mNGF could exert their specific biological role on cultured cells, inducing their peculiar signature pattern of gene expression.14
To further evaluate the synthesis of NGF in the inflamed tissue, we used synovial tissues from adult patients with RA, since synovial tissues were not available from children with JIA (there being no clinical indication for synovial biopsies in JIA). In RA synoviae NGF mRNA expression is extremely high (Figure 2D). Using FLS obtained from synoviae of patients with RA (RA FLS) we found that these cells have a markedly higher expression (p<0.05) of NGF mRNA than fibroblasts obtained from patients with OA (OA FLS), a degenerative joint disease characterised by the absence of chronic inflammation, and control fibroblasts obtained from skin (CTRL FB; Figure 2D). These results clearly indicate that, in the inflamed synoviae, activated fibroblasts and not SFMC are the main NGF producers. Consistent with the findings in JIA synovial fluid (Figure 2A,B) showing proNGF as the predominant form of NGF, we found that RA FLS secreted high levels of proNGF and the amount of proNGF in the conditioned media of RA FLS far exceeded (p<0.001) the concentration of mNGF (Figure 2H).
SFMC p75NTR mRNA levels correlate with disease activity
To evaluate the association of p75NTR mRNA expression with disease severity, we divided patients according to subtypes, with polyarticular JIA being the most severe and oligoarticular JIA the milder disease. We found that SFMC mRNA p75NTR expression levels were highest in patients with polyarticular JIA (n=13) and lowest in patients with persistent oligoarticular JIA (n=29; p<0.01), with patients with extended oligoarticular JIA (n=16) having intermediate levels (Figure 3A). In order to evaluate the association with disease activity at the time of sampling, we analysed the correlation of p75NTR mRNA expression levels with the number of joints of active arthritis, as an estimate of the clinical disease burden, and with C reactive protein (CRP) levels, as a biochemical measure of the degree of inflammation. We found that p75NTR mRNA expression levels in SFMC were significantly correlated with the number of joints with active arthritis at sampling (rs=0.43, p<0.001) (Figure 3B). In order to confirm the difference between patients with a low number of joints with active arthritis and those with a high number of joints with active arthritis, we used a cut-off of 4 joints, which is generally accepted in JIA. We found that patients with ≥5 joints with active arthritis had expression levels of p75NTR (0.053 AU), markedly higher than those of patients with ≤4 joints with active arthritis (0.013 AU). We also found that p75NTR mRNA expressions were correlated with levels of CRP (rs=0.39; p<0.006) (Figure 3C). These results show that high levels of p75NTR mRNA expression at sites of inflammation are associated with the most severe subtype and with clinical and biochemical inflammatory disease activity. Incidentally, we also find in SFMC a significant positive correlation between the p75NTR/TrkA ratio and CRP levels (p=0.01, rS=0.3), erythrocyte sedimentation rate (p=0.04 rS=0.3) and the number of joints with active arthritis at sampling (p=0.03 rS=0.025), suggesting an active role of p75NTR axis during inflammation.
Effects of proNGF on cytokine production by JIA mononuclear cells
High levels of p75NTR and proNGF characterise the site of inflammation in patients with JIA, but our current understanding of the proNGF-p75NTR axis does not offer any data on the possible role of proNGF in the inflammatory response. Since in neuronal cells the binding of proNGF to p75NTR/sortilin appears to induce apoptosis,12 25 we first evaluated whether the addition of proNGF to SFMC affected cell survival. The addition of 200 ng/mL proNGF to SFMC, which are characterised by high p75NTR and low TrkA expression, did not alter the survival of US or LPS-stimulated cells. Similar results were obtained following the addition of 100 ng/mL NGF (figure 4A).

Ex vivo effects of proNGF and NGF on JIA mononuclear cells. (A) No changes in cell viability were observed when mononuclear cells from synovial fluid (SFMC) of patients with JIA were treated for 24 hours with proNGF or mNGF, with or without LPS stimulation. Apoptosis was assessed by Annexin V flow cytometry analysis. The results represent one of three independent experiments performed in duplicate. (B) While proNGF addition alone did not induce the production of interleukin (IL)-6 in unstimulated SFMC, in LPS-activated cells proNGF induced IL-6 release in a dose-dependent manner with a maximal effect at 200 ng/mL (n=4). (C) proNGF stimulation significantly increased IL-6, IL-1β, IL-8 and tumor necrosis factor-α (TNF-α) mRNA expression levels in LPS-stimulated SFMC (n=18) of patients with JIA after 3 hours of incubation, but did not modify IL-10 mRNA expression in either CTRL or JIA mononuclear cells. Mature NGF treatment did not significantly modify proinflammatory cytokine levels or IL-10 mRNA expression in JIA mononuclear cells, while it increased IL-10 mRNA levels in CTRL PBMC. Results are expressed as arbitrary units (AU) and obtained after normalisation with the housekeeping gene GAPDH. (D) proNGF treatment significantly increased IL-6 release after 18 hours (measured by ELISA) in LPS-stimulated SFMC from patients with JIA (n=16) compared with PBMC of healthy CTRL (n=6). Mature NGF treatment did not affect IL-6 protein levels. *p<0.05. JIA, juvenile idiopathic arthritis; LPS, Lipopolysaccharide; NGF, nerve growth factor; PBMC, mononuclear cells isolated from peripheral blood.
We then evaluated whether proNGF affected the expression of inflammatory cytokines in mononuclear cells of patients with JIA and healthy controls. Similar to what we have already demonstrated using mNGF,4 in the absence of TLR4 activation with LPS, the addition of increasing amounts of endotoxin-free proNGF did not induce a significant increase in the release of IL-6 in freshly isolated PBMC of healthy controls (data not shown). There was no dose-dependent induction either in PBMC (data not shown) or in SFMC of patients with JIA (figure 4B), the latter expressing the highest levels of p75NTR, the specific proNGF receptor (see Figure 1A). On the contrary, only when JIA SFMC were costimulated with LPS and proNGF did we observe a dose-dependent increase in the release of IL-6, with a maximal effect at a concentration of 200 ng/mL of proNGF (figure 4B). Interestingly, the effects of mNGF addition to LPS-activated PBMC and SFMC of patients with JIA were different from those of proNGF, as mNGF did not significantly modify IL-6 mRNA expression (figure 4C), an observation that is in line with the previously described inability of mNGF to modulate IL-6 production in JIA mononuclear cells.4 Analysis of IL-6 secreted by SFMC confirmed that the addition of proNGF, but not of mNGF, results in increased IL-6 release (figure 4D). Incidentally, we also found that the addition of proNGF to LPS-activated SFMC induces increased expression of mRNA for IL-1β, IL-8 and TNF-α (figure 4C), an effect that was not observed with the addition of mNGF to LPS-activated SFMC (figure 4C). In these cells, characterised by high p75NTR expression, neither proNGF nor mNGF can upregulate IL-10 mRNA expression, which can instead be significantly induced in control PBMC, expressing high TrkA but low p75NTR, by mNGF but not by proNGF (figure 4C).
To assess whether the effect of proNGF on inflammatory cytokine release is dependent on the binding to p75NTR or to TrkA, SFMCs were incubated with neutralising antibodies against p75NTR or TrkA. In LPS-activated SFMC, the p75NTR blocking resulted in a significant reduction in IL-6 release induced by proNGF (figure 5A). This finding was confirmed using the pharmacological inhibitor LM11A-31, a selective non-peptide ligand of p75NTR, which specifically blocks the binding site of p75NTR for proNGF.26 On the contrary, IL-6 induction by proNGF is not affected when TrkA activity is inhibited with a neutralising antibody (figure 5A), confirming the key role of p75NTR in mediating proNGF activity independently of TrkA.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
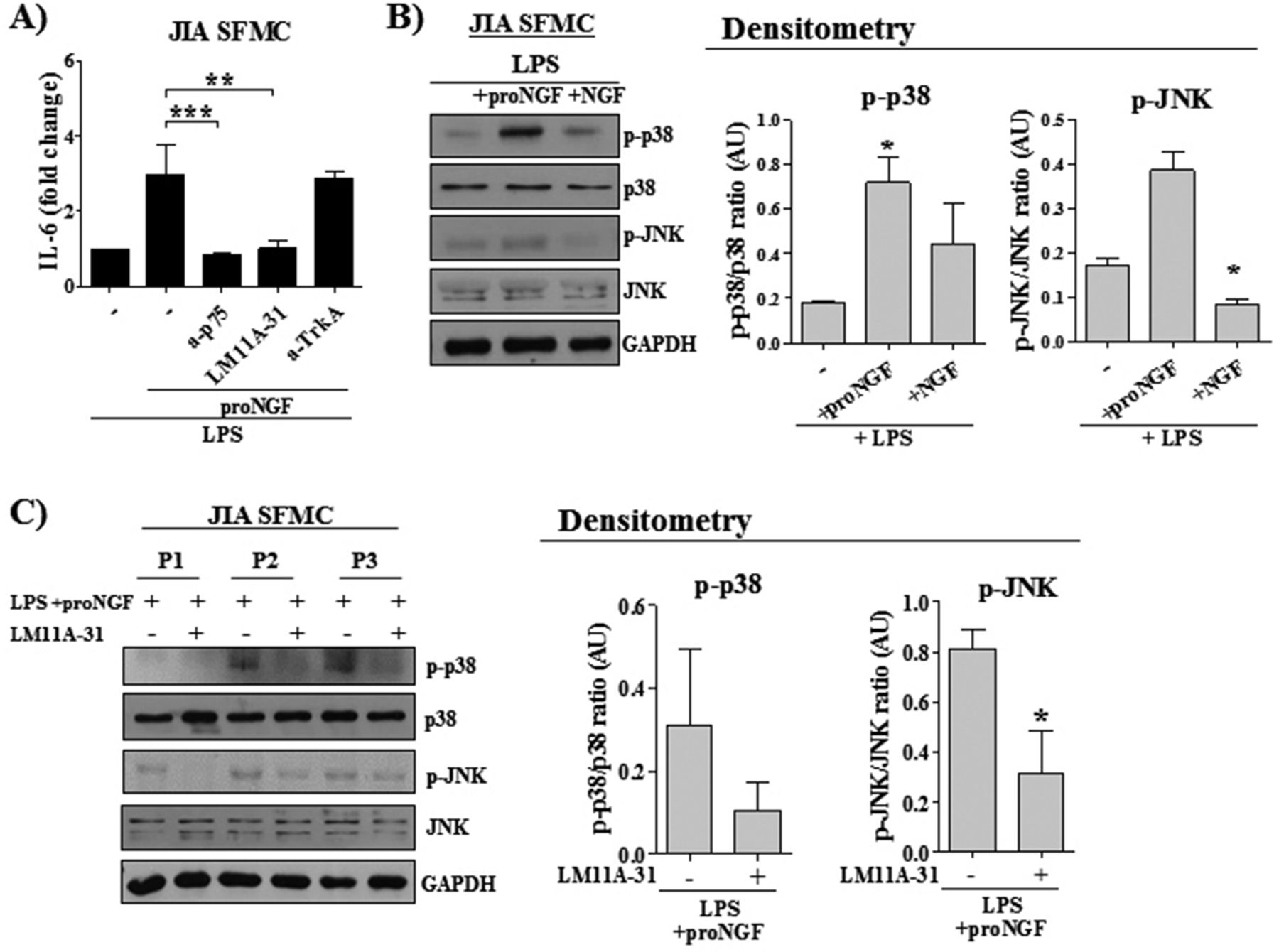
proNGF and NGF activated different intracellular pathways. (A) The neutralisation of p75NTR using either a specific anti-p75NTR antibody (a-p75; 2.5 µg/mL) or using the specific pharmacological inhibitor LM11A.31 (10 nM), decreased interleukin (IL)-6 release in LPS-activated SFMC of patients with JIA (n=16) treated with proNGF. On the contrary, the blocking of TrkA with a neutralising antibody (a-TrkA; 3 µg/mL) did not affect IL-6 release. (B) Western blot shows an increased phosphorylation of p38 and JNK after 5 minutes of proNGF stimulation in LPS-treated SFMC. mNGF treatment did not affect the phosphorylation of these downstream molecules. The result is representative of one out of three independent experiments. Results of the densitometric analysis of all experiments are expressed as arbitrary units (AU). (C) In LPS-activated SFMC of three different patients with JIA, preincubation with the inhibitor LM11A-31 (10 nM), which blocks the binding of proNGF to p75NTR, abolished the phosphorylation of p38 and JNK induced after 5 minutes of proNGF addition. Results of the densitometric analysis of all experiments are expressed as arbitrary units (AU). *p<0.05, **p<0.01, ***p<0.001. JIA, juvenile idiopathic arthritis; LPS, Lipopolysaccharide; NGF, nerve growth factor; SFMC, mononuclear cells isolated from synovial fluids.
Analysis of the pathways involved in p75NTR signalling showed that, in LPS-treated SFMC, the addition of proNGF induced an enhanced phosphorylation of p38 and JNK, which are part of the signalling pathways involved in inflammatory cytokine production (figure 5B). In contrast, the addition of NGF did not induce phosphorylation of p38 or JNK. A reduction in p38 and JNK phosphorylation was observed when the inhibitor LM11A-31 was added together with proNGF, demonstrating that blocking of the binding of proNGF to p75NTR inhibits signalling through classical proinflammatory pathways (figure 5C).
Altogether these results show that in SFMCs, which express high levels of p75NTR, proNGF induces proinflammatory but not anti-inflammatory cytokine expression, and that this effect is mediated through its binding to p75NTR, suggesting that an active proNGF-p75NTR axis may play a proinflammatory role in inflamed joints.
Discussion
Although inflammation has been associated with a marked increase in the synthesis of NGF in tissues1 the effects of mNGF, its precursor proNGF and their receptor signalling in regulating cells and mediators during inflammatory responses are still largely unknown. In this study, we provide evidence that an altered balance between TrkA and p75NTR receptors and an active proNGF-p75NTR axis may contribute to sustain inflammation in chronic arthritis.
We found that mononuclear cells of patients with JIA are characterised by an increased expression of p75NTR and a concomitant decrease in TrkA, at both the mRNA and protein levels, resulting in a highly abnormal ratio between p75NTR and TrkA. The expression of p75NTR in JIA SFMC is 13 times that of TrkA, and even though in these cells there is an increase in TrkA expression compared with JIA PBMC, this is not sufficient to re-establish the balance between the two NGF receptors found in healthy controls, where TrkA is the more expressed receptor. In addition to this abnormal expression of p75NTR, we found in JIA mononuclear cells a parallel increase in the expression of sortilin, the coreceptor of p75NTR to which, as demonstrated in neuronal cells,27 it is structurally and functionally coupled. Since sortilin specifically binds the prosequence of proNGF and increases the affinity of p75NTR for proNGF,12 our results are indicative of the presence of a functional high-affinity proNGF receptor in JIA mononuclear cells.
Previous studies on patients with RA and JIA2 3 and experimental models of arthritis28 29 reported that NGF is enhanced in synovial tissues and fluids. However, the techniques available at the time of these studies were not able to discriminate between the different molecular forms of NGF. Using western blot and newly developed ELISAs,20 we demonstrated in patients with chronic arthritis that proNGF is the highly prevalent form of NGF in the inflamed synovium and that it is actively synthesised by FLS more than by mononuclear cells. The prevalence of proNGF has also been reported in tissues of patients with neurodegenerative diseases, such as Alzheimer’s and Parkinson’s diseases,30 31 which are characterised by local tissue inflammation.32–34 The mechanisms affecting proNGF maturation in inflamed tissues are at present unknown. In the brain of patients with Down syndrome and Alzheimer’s disease, studies by Iulita and Cuello35 showed a deficit in NGF extracellular metabolism that leads to proNGF accumulation. At present, there are no data suggesting that a similar mechanism could occur in peripheral tissues during an inflammatory response and further studies are needed.
The presence of high levels of proNGF in peripheral tissues15 20 36 has recently been the focus of a number of studies since it is becoming clear that proNGF and mNGF have different, even opposite, effects on neuronal cells14 37 and, as recently reported, also on macrophages.38
To characterise the effect of an active p75NTR axis on inflammation, we treated JIA SFMC ex vivo with either proNGF or mNGF. While in neurons the binding of proNGF to p75NTR/sortilin leads to apoptosis,27 incubation of JIA cells with either proNGF or mNGF did not lead to changes in cell viability. In the absence of TLR stimulation, the addition of proNGF alone did not induce a significant increase in the expression of inflammatory cytokines in mononuclear cells of either patients with JIA or healthy controls, regardless of their level of p75NTR expression. In contrast, the addition of proNGF to LPS-activated SFMC leads to an increase in the expression of proinflammatory cytokines, such as IL-1β, IL-6, IL-8 and TNF-α, while mNGF addition did not induce inflammatory cytokine expression. The addition of proNGF to LPS-activated SFMC did not modify the expression of IL-10, an anti-inflammatory cytokine whose synthesis is instead specifically induced by mNGF in monocytes4 and in control PBMC, both cell types characterised by high expression levels of TrkA. Consistent with this finding, the incubation of LPS-activated SFMC with proNGF significantly increased the phosphorylation of p38 and JNK, activating pathways involved in inducing inflammatory cytokine expression,39 while the addition of mNGF to LPS-activated SFMC did not have these effects. Therefore, in line with recent data on neurons40 and on PC12 cells,14 the biological effects and the signalling pathways activated in JIA cells by proNGF differ from those of mNGF. These data indicate that the binding of proNGF to p75NTR, but not of mNGF, is specifically involved in the induction of inflammatory cytokine expression. Confirming this hypothesis, we found that the blocking of p75NTR with a neutralising antibody or LM11A-31, a selective non-peptide ligand of p75NTR that specifically blocks the binding site of p75NTR for proNGF,26 specifically inhibited phosphorylation of p38 and JNK and abolished the IL-6 production induced by proNGF, while the inhibition of TrkA did not affect IL-6 levels. A similar inhibitory effect of LM11A-31 on JNK phosphorylation has been described after LM11A-31 blocking of p75NTR in hippocampal neurons in an in vitro model of Alzheimer’s neurodegeneration.41 The functional improvement in motor function and coordination observed in a mouse model of spinal contusion injury after in vivo administration of LM11A-31 has been associated with a significant reduction in JNK activity in the spinal cord, further highlighting the central role of this pathway in proNGF/p75 NTR signalling.42
The molecular basis of the different, even opposite, effects of mNGF and proNGF that use the same receptor systems is not yet completely clarified. The most reasonable explanation involves the different binding affinity of the two NGF forms, with mNGF showing the higher affinity for TrkA and proNGF for the p75NTR/sortilin complex.9 In our previous studies on human mononuclear cells and monocytes of healthy donors, expressing significantly more TrkA than p75NTR, the binding of mNGF to TrkA reduced inflammatory cytokine production while increasing the release of anti-inflammatory cytokines. These effects on cytokine expression were associated with the inhibition of nuclear NF-κB translocation, inhibition of GSK3 activity and induction of Akt.4 The anti-inflammatory effects of NGF on cytokine expression and on intracellular signalling pathways were abolished by blocking TrkA, but not p75NTR.4 Interestingly, Jiang and coworkers43 have reported that also mNGF, through binding to p75NTR in human dendritic cells that express p75NTR but not TrkA, induced cytokine expression and dendritic cell maturation by specifically activating p38 and NF-κB pathways. Altogether, these data suggest that, more than the relative concentrations of the two ligands (ie, mature and proNGF), it is the relative levels of expression of the two receptor systems (ie, TrkA and p75NTR/sortilin) that direct the immune cells towards a proinflammatory or an anti-inflammatory response. In a study using the PC12 neuronal cell line, it was shown that proNGF can have neurotrophic activity in primed PC12 or apoptotic activity in unprimed PC12, two cell conditions characterised by a different ratio of TrkA and p75NTR, further suggesting that the biological activity of proNGF is dependent on the relative expression levels of TrkA and p75NTR.44
In agreement with this hypothesis are our data on the differential effects of proNGF in LPS-activated cells on cytokine production determined by p75NTR expression levels. Indeed, the addition of proNGF to healthy donor cells, with higher TrkA than p75NTR expression, did not enhance inflammatory cytokine expression. In contrast, the addition of proNGF to JIA SFMC expressing higher levels of p75NTR led to enhanced production of inflammatory cytokines. It is therefore highly conceivable that the high expression of p75NTR in immune cells coupled with high levels of proNGF, released by fibroblasts in inflamed synovium, may contribute to amplify the inflammatory response in chronic arthritis. It is worth noting that data consistent with this hypothesis have been reported in diabetic retinopathy. In human and rat retinas, high expression levels of p75NTR and decreased phosphorylation of TrkA are associated with a shift from mNGF to the immature proNGF form.45 46 Moreover, studies in experimental diabetes using p75NTR knockout mice have shown a reduced inflammatory response and a decrease in TNF-α release in the retina with a beneficial effect on glial activation and ganglion cell loss,47 further supporting the proinflammatory role of p75NTR.
An additional, although indirect, indication of the proinflammatory role of p75NTR was obtained from the analysis of p75NTR expression in patients with JIA divided into groups according to their subtype. We found that the levels of p75NTR expression are correlated with disease severity and activity as documented by the direct correlation with JIA subtypes and with the number of active joints and CRP levels. Altogether these data point to a proinflammatory action of proNGF which is mediated through its binding to p75NTR and not through TrkA.
In conclusion, this study provides new evidence for a role of p75NTR and its preferential ligand proNGF in amplifying inflammatory responses in chronic arthritides. The relative expressions of p75NTR and TrkA, as well as the relative concentrations of proNGF and mNGF forms, may regulate inflammatory responses and may represent an additional mechanism in the progression of inflammatory joint disease, with high levels of proNGF and high expression of p75NTR tilting the balance towards proinflammatory pathways. Inhibition of the p75NTR-proNGF axis and its effects on the proinflammatory loop should be investigated as a possible new therapeutic approach in chronic arthritis and in animal models. In this respect, the in vivo efficacy of LM11A-31, the non-peptide ligand that blocks the binding site of p75NTR for proNGF, has recently been demonstrated in animal models of spinal cord injury42 and Alzheimer’s disease.48 A clinical trial (NCT03069014) assessing the safety of LM11A-31-administration in patients with Alzheimer’s disease is currently ongoing.
Acknowledgments
The authors wish to thank Dr Marcello Allegretti and Dr Candida Cesta of Dompè Farmaceutici SpA for kindly providing p75NTR antagonists, Professor Maria Egle De Stefano for helpful discussion during manuscript preparation. The study was founded by Ospedale Pediatrico Bambino Gesù’ "Ricerca Corrente" to FDB.
References
Footnotes
Contributors GM performed in vitro experiments. MS performed ELISAs. IC and GP performed RT-PCR. LM performed WB analysis. SMM collected samples and clinical data from patients. DPM conducted statistical analysis. AM isolated synoviocytes. FDB and LBL planned the experiments. GM, FDB and LBL wrote the manuscript.
Competing interests None declared.
Patient consent Parental/guardian consent obtained.
Ethics approval The study was approved by the Ethical Committee of the Bambino Gesù Children’s Hospital and written consent was obtained from parents of children as appropriate.
Provenance and peer review Not commissioned; externally peer reviewed.