Article Text

Abstract
Background The DNA of rheumatoid arthritis synovial fibroblasts (RASF) is globally hypomethylated; this contributes to an aggressive behaviour. In an attempt to remethylate these cells, we supplemented with methyl donors. We investigated the possible interference of microRNAs (miRs).
Material and methods RASF were treated with L-methionine or betaine. Transcripts of de novo methyltransferases (DNMTs) and miRs were measured by real-time PCR, and a transcription PCR array was performed. Levels of homocysteine, matrix metalloproteinase-1 (MMP-1) and global DNA methylation were determined. Transfection with lipofectamine was performed with specific pre-miRs and anti-miRs, such as miR29 and let7f.
Results L-methionine was more efficient to increase DNA methylation than betaine. This was associated with a reduced expression of DNMT3A mRNA in betaine-treated RASF. Betaine increases the expression of miR29 in RASF which targets DNMT3A, thereby limiting the remethylation process. Nevertheless, betaine inhibited the expression of multiple transcription factors, decreased the release of MMP-1, biosynthesis of homocysteine and cell migration.
Conclusion Alterations in cellular miRs profiles, in particular the upregulation of miR29, which targets DNMT3A, may limit the efficiency of betaine if it is used as DNA remethylating agent. However, L-methionine also has similar impact on miR29 expression. On the other hand, betaine has multiple other beneficial effects on the activated phenotype of RASF; it is not excluded that the effect of betaine on DNMT3A is, at least in part, indirect. Clinical trials with betaine could be promising.
- Rheumatoid Arthritis
- Synovitis
- Pharmacogenetics
- Fibroblasts
This is an Open Access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/
Statistics from Altmetric.com
Key messages
Rheumatoid arthritis synovial fibroblasts (RASF) showed global DNA hypomethylation. Treatment with methyl donors could be limited by the intrinsically activated recycling of polyamines that compete for S-adenosyl methionine (SAM).
We showed here that a microRNAs-dependent mechanism selectively target DNA methylation possibly limiting the remethylation process. On the other hand, betaine showed multiple beneficial effects unrelated to DNA methylation, for example, inhibiting the expression of multiple transcription factors.
Large interindividual variations can be expected in outcomes of treatments with methyl donors, depending on the active mechanisms limiting the remethylation process. Nevertheless, clinical trials with betaine could be promising.
Introduction
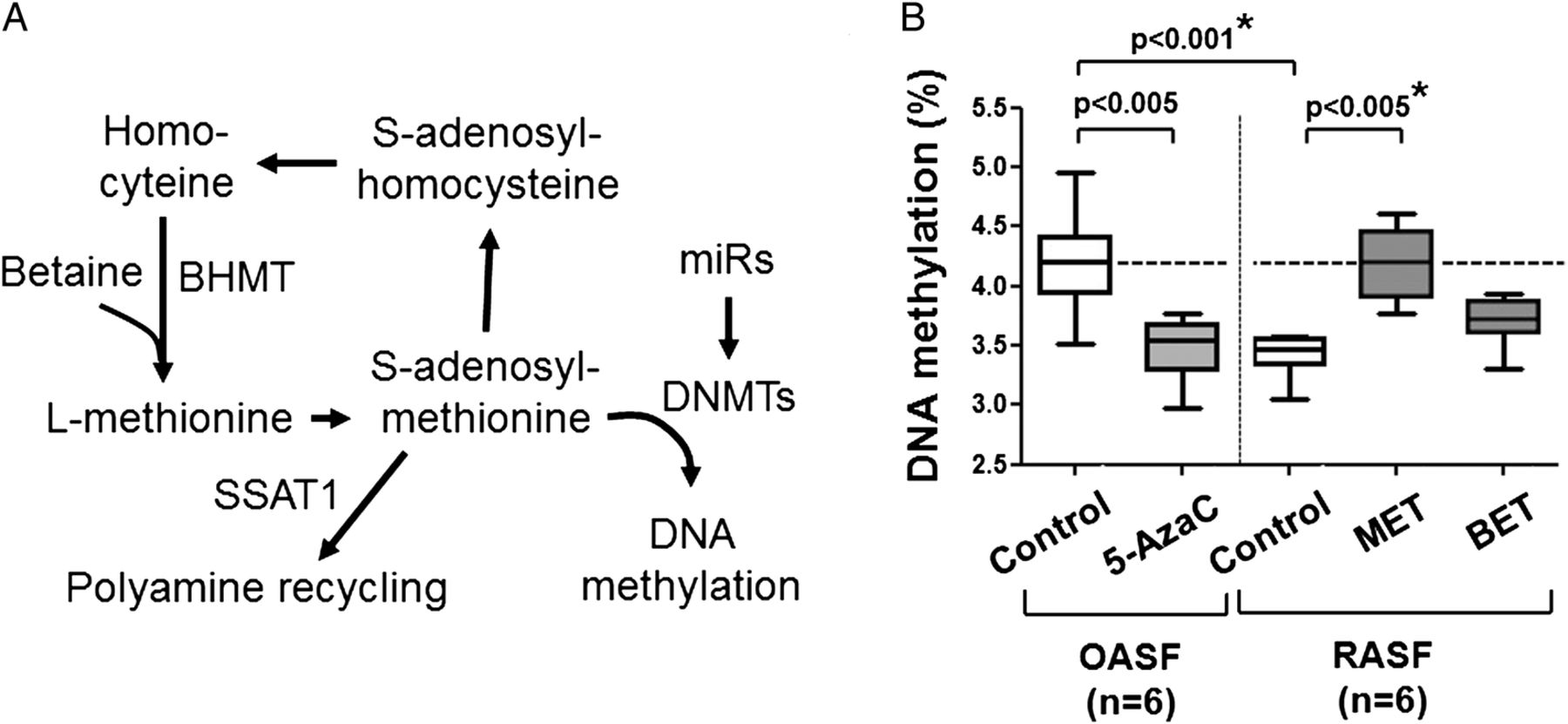
The global DNA hypomethylation in rheumatoid arthritis synovial fibroblasts (RASF) is associated with an intrinsically activated and aggressive phenotype.1 DNA methylation is performed by maintenance methyltransferase (DNMT1) and de novo methyltransferases (DNMT3A and DNMT3B). These enzymes catalyse a transmethylation reaction using S-adenosyl methionine (SAM) (figure 1A). L-methionine and betaine feed into the methionine cycle as precursors of SAM and provide their methyl group.2 ,3 Betaine is involved in the remethylation of homocysteine to synthesise L-methionine, a pathway catalysed by betaine-homocysteine methyltransferase (BHMT).4 L-methionine and betaine might represent strategies to reverse DNA hypomethylation locally in rheumatoid arthritis (RA) synovial tissue and systemically in peripheral T lymphocytes of patients with RA.5 However, in RASF, the polyamine recycling pathway excessively consumes SAM, due to an increased expression of spermine/spermidine N1-acetyltransferase (SSAT1), thereby interfering with the DNA methylation process.6 ,7 We show here that, in comparison with L-methionine, the efficiency of betaine to remethylate RASF is lower, and this might be due to the induction of microRNAs (miRs) targeting the expression of DNMTs, for example, miR9,8 miR299 ,10 and let7f.11 ,12 We explored how miRs may selectively target mechanism of DNA methylation. On the other hand, we also show that betaine induced several beneficial changes unrelated to DNA methylation.

(A) One-carbon cycle and polyamine metabolism regulate DNA methylation. (B) RASF showed decreased global DNA methylation that is comparable to OASF on 5-azacytidine. Treatment of RASF with L-methionine restored DNA methylation; betaine was less efficient (2 weeks, 20 mM). Statistics: Box-and-Whisker plots, Mann-Whitney U-test between OASF and RASF, Wilcoxon signed rank test between OASF before and after treatment with 5-azacytidine, as well as RASF before and after treatment with L-methionine, *confirmed by Kruskal-Wallis with Dunn's post hoc tests (overall: p<0.01; *confirmed for control OASF vs control RASF, as well as between control RASF and RASF treated with L-methionine, n=6). 5-AzaC, 5-azacytidine; BET, betaine; BHMT, betaine-homocysteine methyltransferase; DNMT, de novo methyltransferase; MET, L-methionine; miRs, microRNAs; OASF, osteoarthritis synovial fibroblasts; RASF, rheumatoid arthritis synovial fibroblasts; SAH, S-adenosyl homocysteine; SAM, S-adenosyl methionine; SSAT, spermine/spermidine N1-acetyltransferase.
Materials and methods
Cell cultures
RASF (n=6–9) and osteoarthritis synovial fibroblasts (OASF) (n=6) were obtained after joint replacement (Schulthess Clinic, Zurich, Switzerland). Patients fulfilled the diagnostic criteria for RA13 or osteoarthritis (OA).14 The procedure was approved by the Ethics Committee of the University Hospital Zurich, and canton of Zurich, Switzerland. SW982 cells from synovial sarcoma and HepG2 from hepatocellular carcinoma were obtained from American Type Culture Collection (ATCC)/Laboratory of the Government Chemist (LGC) Standards GmbH. RASFs were treated with L-methionine or betaine (0–50 mM, Sigma-Aldrich). The medium was replaced once a week with the same concentration of the methyl donors. After 2 weeks, cell culture supernatants were collected and cells were detached using Accutase (PAA Laboratory). OASF were passaged and treated for 2 weeks with 5-azacytidine (0.1 μM, Sigma-Aldrich), replaced in fresh medium twice a week. Proliferation rates were measured using a Casy (Innovatis) cell counter and the xCELLigence system (ACEA Biosciences). The scratch/migration assay was performed as previously described.15
Real-time PCR
RNA was isolated using ReliaPrep minicolumn kit (for mRNA, Promega) or miRNeasy kit (for total RNA, Qiagen) and stored at −20°C. Genomic DNA was isolated using the QIAamp DNA kit (Qiagen). Qubit 2.0 Fluorometer (Invitrogen) was used to quantify the amounts of nucleic acids. Reverse transcription was performed using SuperScript First-Strand Synthesis System (Invitrogen) or MultiScribe Reverse Transcriptase (Life Technologies). The following primers were used for quantitative real-time PCR: DNMT1 fwd 5′-TAACAGAAAAGGAATGTGTGAAGG-3′, rev 5′-TATTTCTGTTTGCAGAAATTCGTCC-3′; DNMT3A fwd 5′-TATTGATGAGCGCACAAGAGAGC-3′, rev 5′-GGGTGTTCCAGGGTAACATTGAG-3′; DNMT3B fwd 5′-TGGCGATGGCAAGTTCTCCGAG-3′, rev 5′-CAGTGCCACCAGTTTGTCTGC-3′. A commercially available TaqMan MicroRNA Assay (Life Technologies) was used to measure miRs (miR9, miR29, miR203, let7f). PCR was conducted using Taq PCRx DNA Polymerase and a 7900 HT Fast Real-time PCR System (Life Technologies). Regarding miRs, PCR was performed using no AmpErase UNG, and single tube TaqMan microRNA assay (Life Technologies). HPRT1 was used as an endogenous control for normalisation and for miRs mean values of RNU6B and RNU48 endogenous controls were used. In addition, Transcription Factors RT2 Profiler PCR Array (Qiagen) was performed.
Enzyme linked immunosorbent assays
Global DNA methylation was quantified using the 5-mC DNA ELISA kit (Zymo Research). SSAT1 levels in cell homogenates were measured using a SAT1 ELISA kit (MyBioSource). L-homocysteine was quantified using the Axis Homocysteine Enzyme Immunoassay kit. DuoSet ELISA Development kit (R&D systems) was used to measure matrix metalloproteinase-1 (MMP-1).
SSAT1 enzymatic activity assay
The assay was derived from the method of Lin et al.16 Adherent cells were lysed using 500 µL Sigma CelLytic M solution. The homogenate was added with 500 mL of 50 mM Tris-HCl, 1 mM EDTA, pH 7.5 (all from Sigma-Aldrich). It was adjusted to 100 µg/mL protein using a bicinchoninic acid (BCA) protein assay (the homogenate can be diluted using 25 mM tris-HCl and 0.5 mM EDTA, pH 7.5). This adjusted homogenate was used for SSAT1 ELISA. The SSAT1 activity assay sample mixture consisted of 196 µL sample and 4 µL 25 mM acetyl-coenzyme A (end concentration 0.5 mM (Sigma-Aldrich)). To 25 µL of spermine solution (of different concentrations between 1 and 4 mM (Sigma-Aldrich), prepared in 25 mM Tris-HCl and 0.5 mM EDTA, pH 7.5) 25 mL sample mixture was added. To a final volume of 100 mL 50 mL Ellman's reagent (2 mM DTNB, 1 mM EDTA, 50 mM Tris-HCl, pH 7.5 (Sigma-Aldrich)) was added. The sample mixtures were then incubated for 5, 10, 15 or 20 min at 37°C. After heating at 65°C for 5 min, the absorption was measured at 450 nm. Bioactive human SSAT1 protein purified from a HEK293T cells SSAT1 overexpression lysate (OriGene) was used as standard.
Flow cytometry
Apoptosis was measured using Phycoerythrin Annexin V Apoptosis Detection Kit I (BD Pharmingen). For comparison, apoptosis was induced in RASF within 24 hours using 5 μM cycloheximide (Sigma-Aldrich). Intracellular staining was performed using BD Perm2 solution (Becton-Dickinson), as well as specific primary antibodies: anti-BHMT and anti-SSAT1 or isotype control antibodies (Abcam). The labelling was revealed using fluorescein-conjugated secondary antibodies (Jackson Laboratory).
Transfection
Transfection was conducted using Lipofectamine 2000 (Invitrogen), negative control Pre-miR, Pre-miR29, Pre-miR-7f (50 nM) and negative control Anti-miR, Anti-miR29 and Anti-miR-7f (100 nM) (Ambion Pre-miR miRNA Precursors and Ambion Anti-miR miRNA Inhibitors, respectively).
Statistics
Mann-Whitney U-test, Wilcoxon signed rank test, Kruskal-Wallis with Dunn's post hoc tests and Spearman's correlation were used to reveal the levels of significance and the preliminary relation among variables. Data are mainly presented as Box-and-Whisker plots showing the median (horizontal line), 50% IQR (Box, IQR) and up to 1.5 times IQR (Whiskers).
Results
Methyl donors restores global DNA methylation in RASF
We measured cell proliferation and apoptosis with different concentrations of L-methionine and betaine; 50 mM betaine was cytostatic, but not apoptotic (see online supplementary figure S1A–C). In RASF and SW982 cells, which express BHMT, betaine was cytostatic, but not in HepG2 cells, which lack functional BHMT (see online supplementary figure S1D). In all the follow-up experiments, we used concentrations of ≤20 mM, which are neither cytostatic nor apoptotic. We confirmed that RASF showed decreased global DNA hypomethylation that was comparable to OASF treated with a low non-toxic dose of 5-azacytidine (figure 1B). Treatment of RASF with L-methionine or betaine restored DNA methylation; however, L-methionine was significantly more efficient than betaine (figure 1B). L-methionine and betaine restore DNA methylation, but not at the same degree at a given molar concentration.
supplementary figures
In RASF, considering that a baseline DNA methylation of 3.5%, an increase to a ‘normal’ value of 4.2%, as it is achieved with L-methionine, would corresponds to (95% CI ) a 17.2% to 26.8% increase of methylation of the whole DNA. For betaine, the 95% CI was 3.4% to 6.6%; this would correspond to a Z-value of 10.4 and a highly significant difference between L-methionine and betaine.
SSAT1 levels and DNA methylation on methyl donors
We were interested to identify the pathway which is altered by methyl donors. Previously, we have shown that the global DNA hypomethylation in RASF is associated with higher expression of SSAT1, that is, an enhanced recycling of polyamines. However, the increases in DNA methylation on methyl donors were not significantly associated with changes in this pathway (see online supplementary figure S2A,B). In order to validate the flow cytometric measurement, different methods to assess SSAT1 levels and activity were compared (see online supplementary figure S3A,B). As expected, the three methods showed highly significant correlations. However, it is important to note that SSAT1 activity can vary depending on the intracellular conditions. For example (see online supplementary figure S3C), using a colorimetric assay and a Lineweaver-Burk double reciprocal plot, we compared SSAT1 activity in protein extracts from OASF and RASF (n=3 each). In RASF, Km was lower (0.33 mM in RASF, compared with 0.52 mM in OASF) and Vmax was either similar or lower (0.012–0.020 mM/min in RASF, compared with 0.019 mM/min in OASF). Thus, the activity of SSAT1 in RASF appeared limited by competitive (in 1/3 RASF) and/or mixed intracellular inhibitions (in 2/3 RASF), more than in OASF.
DNMT3A expression is more critical than SSAT1 on betaine treatment
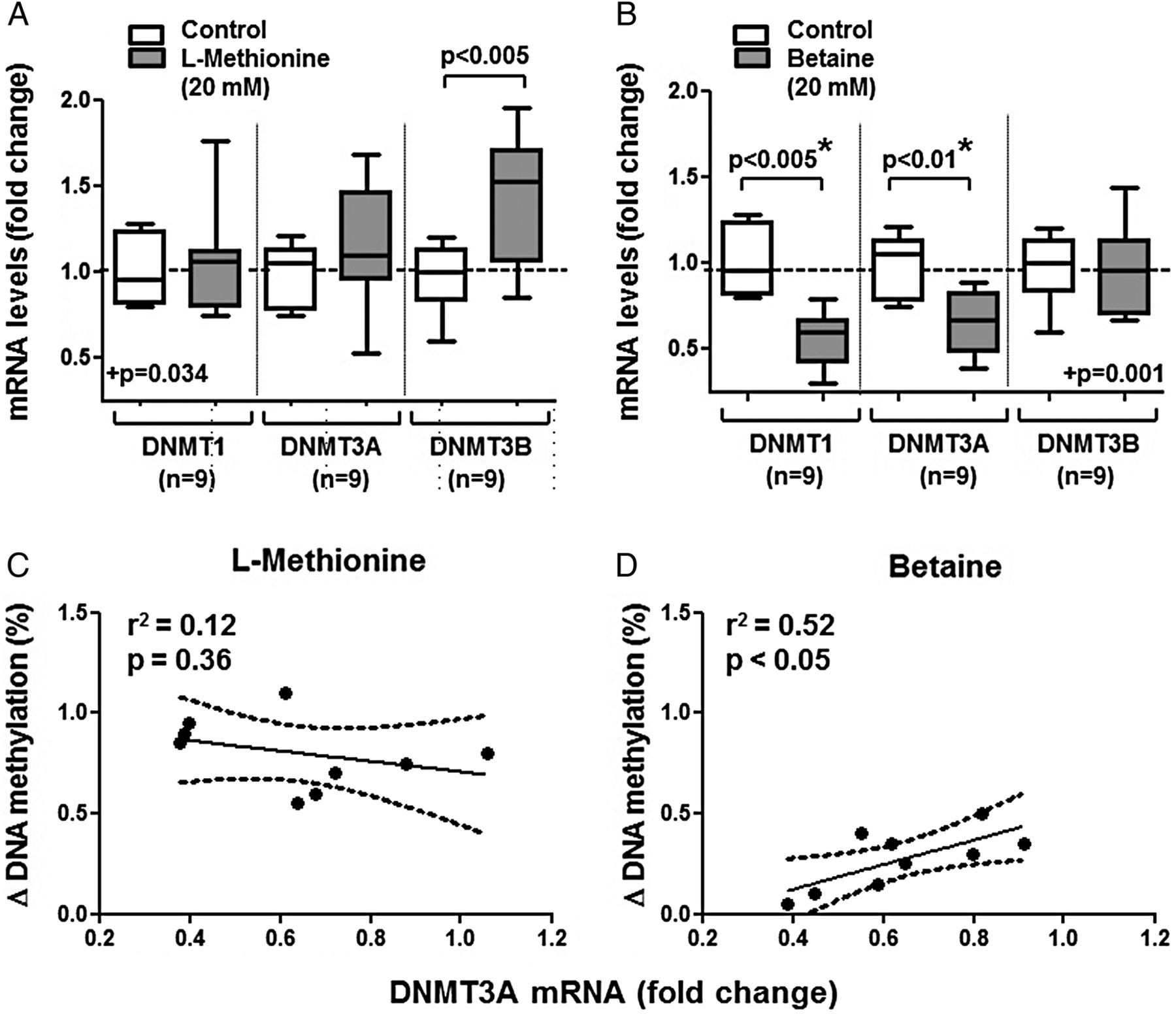
In comparison to untreated RASF, changes in DNMT transcripts occurred after treatment with methyl donors (figure 2A,B); in cells treated with betaine, but not in those treated with L-methionine, DNMT1 and DNMT3A mRNAs were reduced by 35% and 40%, respectively. A significant association between increases in DNA methylation and changes in DNMT3A expression was observed on betaine, but not on L-methionine (figure 2C,D). Interestingly, DNMT3B transcripts were increased on L-methionine (figure 2A); however, no positive association between changes in DNA methylation and DNMT3B transcripts was found (data not shown). In contrast, DNMT1 transcripts showed no change on L-methionine and also no positive association with changes in DNA methylation.

(A and B) Changes in the levels of DNA methyltransferases (DNMTs) transcripts on treatment of RASF for 2 weeks with 20 mM L-methionine (A) or betaine (B). The L-methionine-treated cells showed a trend towards more DNMT3B mRNA. At opposite, on betaine (but not L-methionine), expressions of DNMT1 and DNMT3A mRNAs were reduced by 35% and 40%. (C and D) A significant association between increases in DNA methylation and changes in DNMT3A transcripts expression was observed after betaine (D), but not after L-methionine-treated treatment (C). Statistics: (A and B) Box-and-Whisker plots, Wilcoxon signed rank test between RASF before and after treatment with L-methionine or betaine, *confirmed by Kruskal-Wallis with Dunn's post hoc tests (overall: p<0.05 for A. and p<0.001 for (B), *separately confirmed in (B) regarding DNMT1 and DNMT3A, n=9); (C and D) linear regression lines with 95% CI, Spearman's rank correlation coefficients. DNMT, de novo methyltransferase; RASF, rheumatoid arthritis synovial fibroblasts.
Increased miR29 expression might limit DNA remethylation in RASF
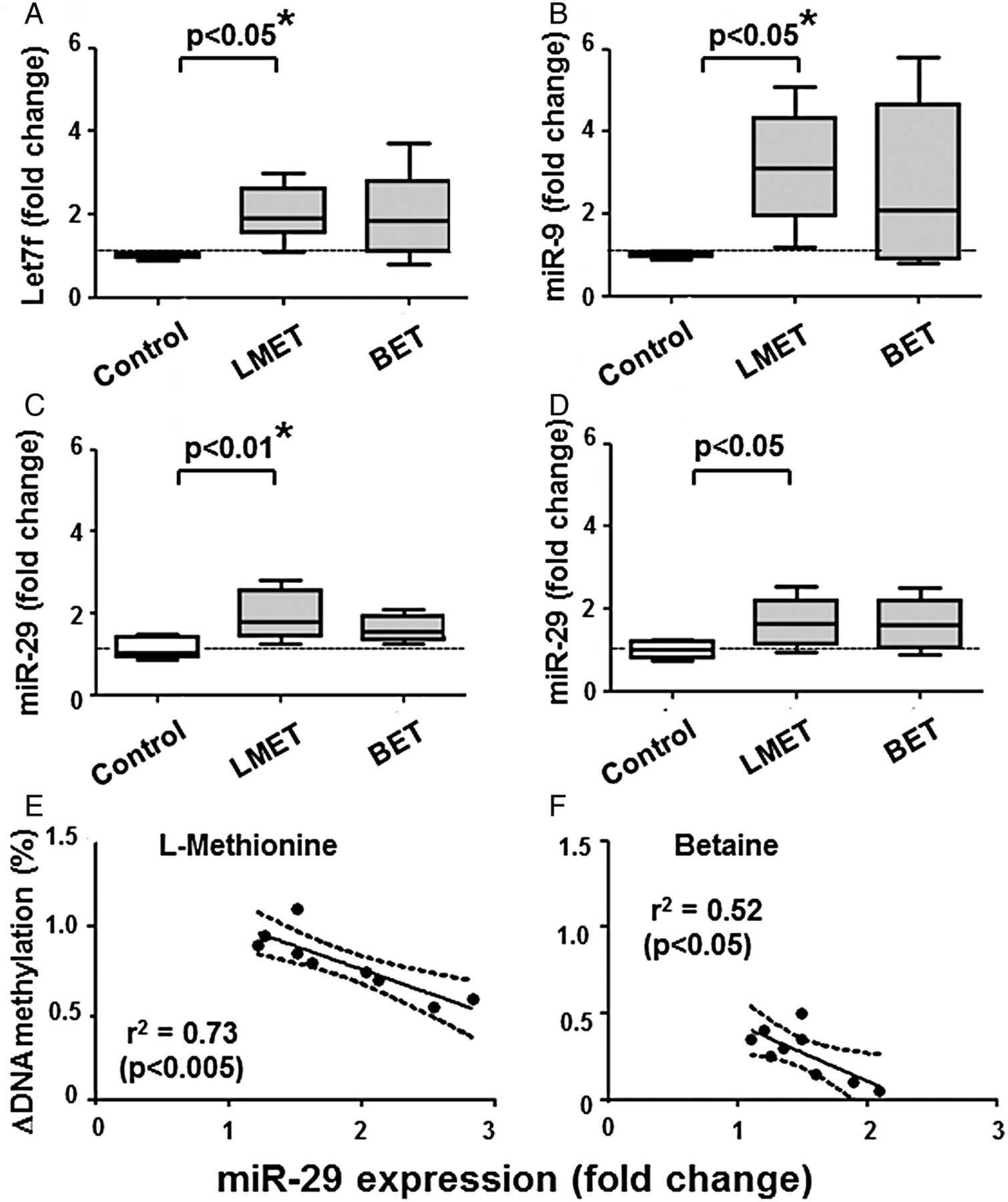
We measured the levels of miRs described to have DNMTs as targets (ie, miR9, miR29 and let7f), as well as miR203 with promoter hypomethylated in RASF. Indeed, we found that the level of some of these miRs changed on L-methionine (figure 3A–D), also according to the 95% CI. Thus, L-methionine increased the levels of miR29 (95% CI 1.14-fold to 1.66-fold change, n=9), miR203 (1.34 to 1.86), miR9 (1.54 to 5.46) and let7f (1.67 to 2.33). Changes in DNA methylation on methyl donors inversely correlated with increases in miR29 expression (figure 3E), but not with the other miRs tested. The regression curves obtained on L-methionine or betaine have similar slopes, but the change in DNA methylation was lower in cells treated with betaine.
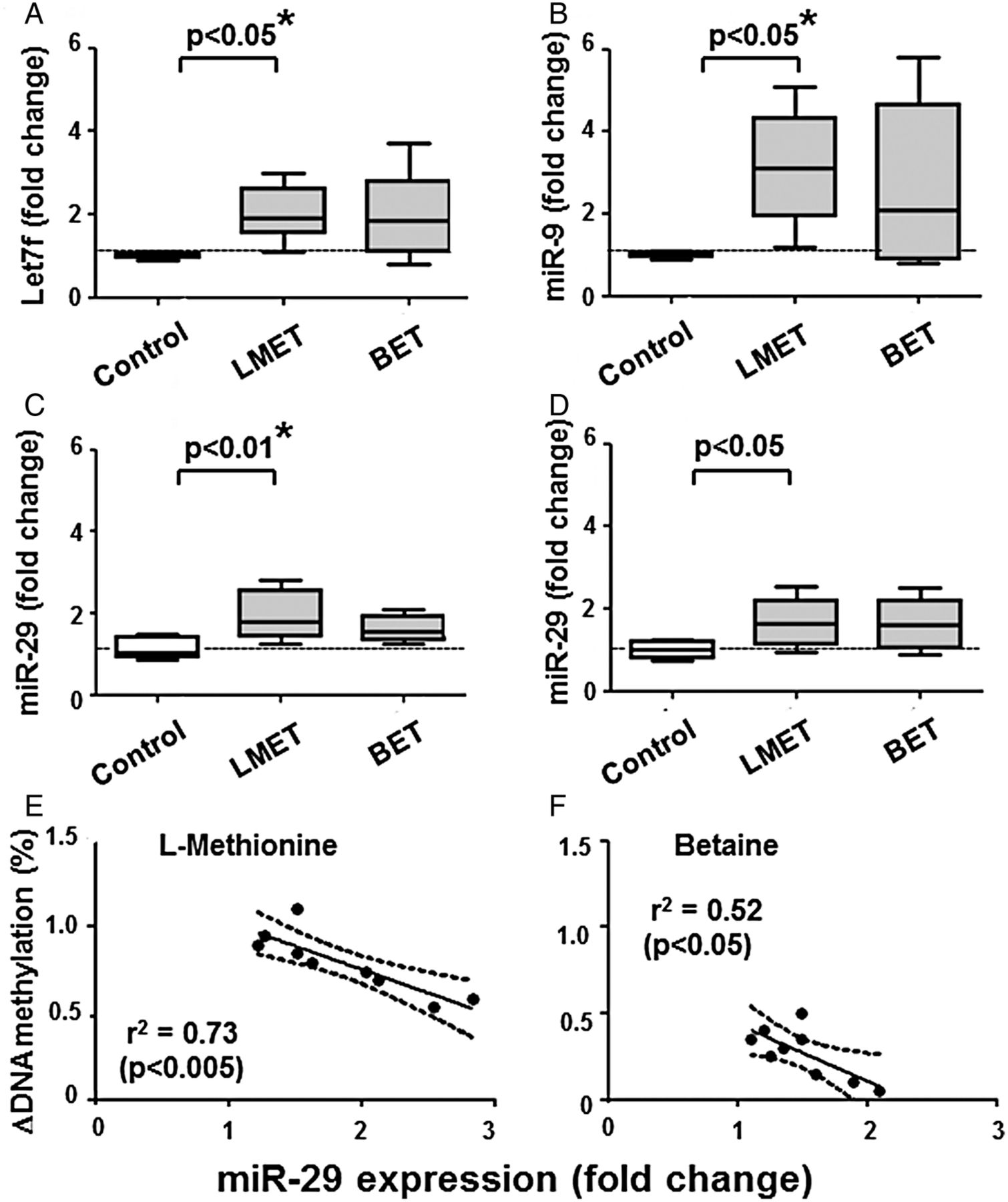
Changes in microRNA expression on treatment of RASF for 2 weeks with 20 mM L-methionine or betaine. (A and B) let7f and miR9 increased significantly in L-methionine-treated cells. (C and D) miR29 and miR203 increased after both treatments, but only miR29 on L-methionine reached the level of significance after analysis of variance. (E and F) Changes in DNA methylation on L-methionine (E) or betaine (F) inversely correlated with increases in miR29 expression. This relationship is observed in spite that L-methionine and betaine have different efficiencies to increase DNA methylation in RASF. Statistics: (A–D). Box-and-Whisker plots, Wilcoxon signed rank test between RASF before and after treatment with L-methionine or betaine, *confirmed by Kruskal-Wallis with Dunn's post hoc tests (overall: p<0.005 for (A—C), p=0.061 for (D); * separately confirmed in (A–C) regarding let7f, miR9 and miR29 on L-methionine); (E and F) linear regression lines with 95% CI, Spearman's rank correlation coefficients. BET, betaine; MET, L-methionine; miRs, microRNAs; RASF, rheumatoid arthritis synovial fibroblasts.
mir29 targets DNMT3A in synovial fibroblasts
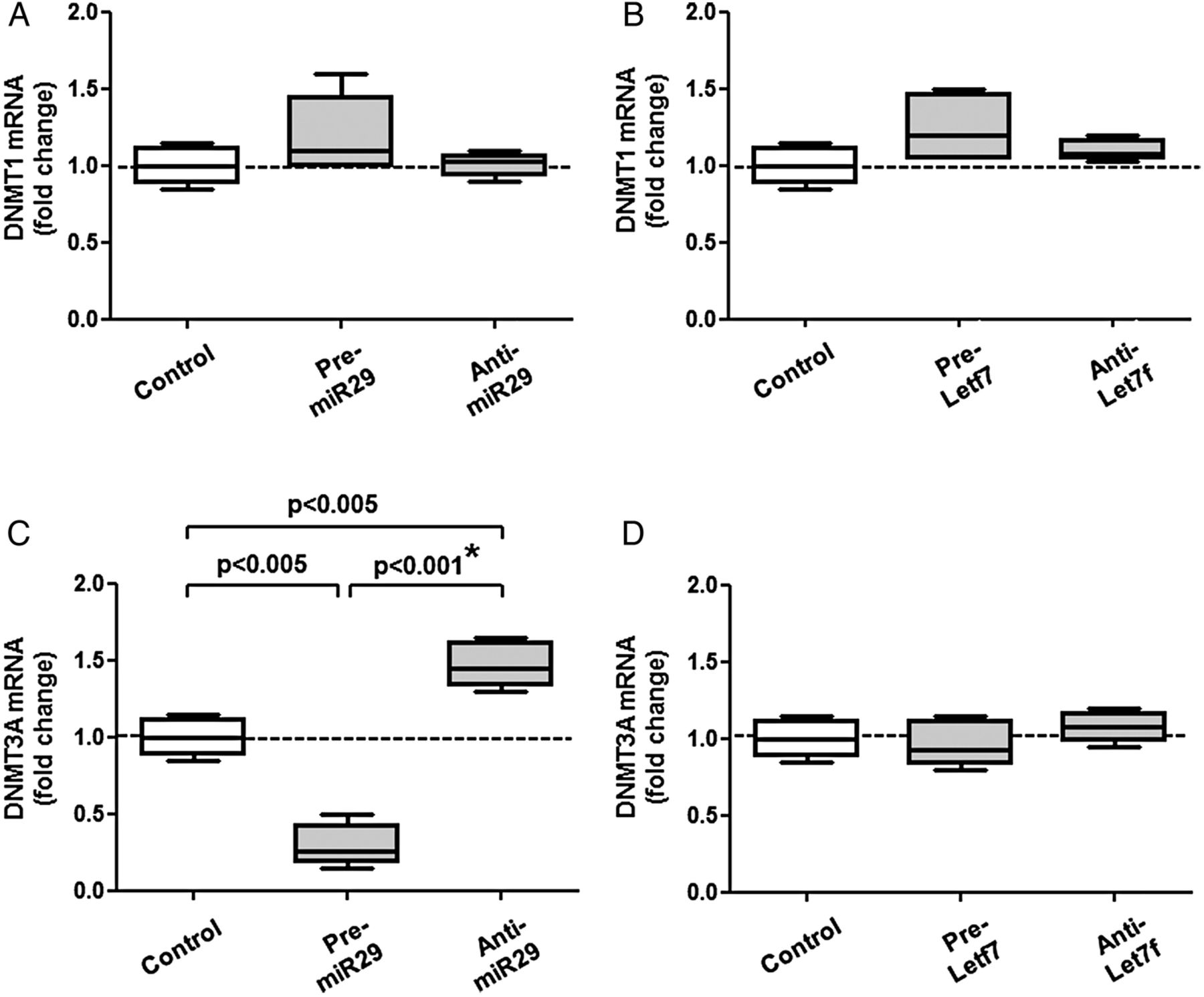
We confirmed that DNMT3A is a direct target of miR29. Pre-miR29 or pre-let7f did not affect the expression of DNMT1 transcripts (figure 4A,B). However, pre-miR29—but not pre-let7f—reduced DNMT3A transcripts by 70% (figure 4C,D). The relationship between miR29 and DNMT3A transcripts was also shown using anti-miRs (figure 4A,D); the difference between pre-miR29 and anti-miR29 was highly significant and confirmed by the post hoc corrected analysis of variance. Finally, we tested the hypothesis that anti-miR29 restores the level of DNMT3A mRNA in the presence of methyl donors (figure 5A,B). Indeed, we observed a 138+43% increase of DNMT3A on anti-miR29 transfection on betaine. The differences between betaine alone and betaine plus anti-miR29 for DNMT1 and DNMT3A transcripts were significant and confirmed by the post hoc corrected analysis of variance.
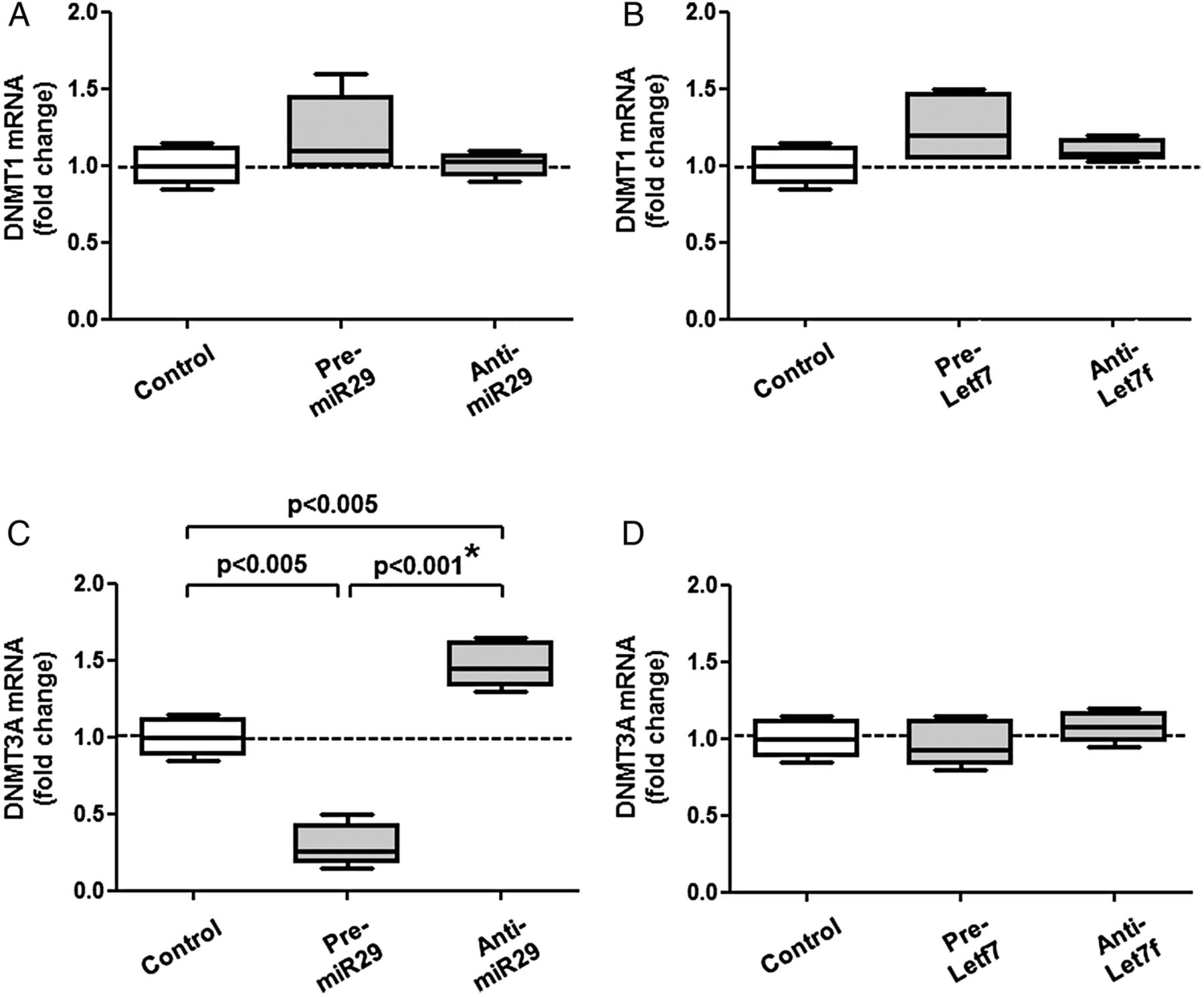
(A and B) Pre-miR29/anti-miR29 and pre-let7f/anti-let7f did not affect the expression of DNMT1 transcripts. (C and D) Pre-miR29 decreased DNMT3A mRNA, while anti-miR29 increased it, suggesting that DNMT3A transcripts are direct targets of miR29 in RASF (C); the difference between pre- and anti-miR29 was considered as significant after analysis of variance. For comparison (D), pre-let7f/anti-let7f had much less effects on DNMT3A mRNA. Statistics: Box-and-Whisker plots, Wilcoxon signed rank test between RASF before and after transfection with pre-miR and anti-miR, *confirmed by Kruskal-Wallis with Dunn's post hoc tests (overall: p=0.28 for (A), p=0.11 for (B), p<0.005 for (C), p=0.25 for (D); * confirmed separately for DNMT3A on pre-miR29 and anti-miR29, n=6). DNMT, de novo methyltransferase; miRs, microRNAs; RASF, rheumatoid arthritis synovial fibroblasts.
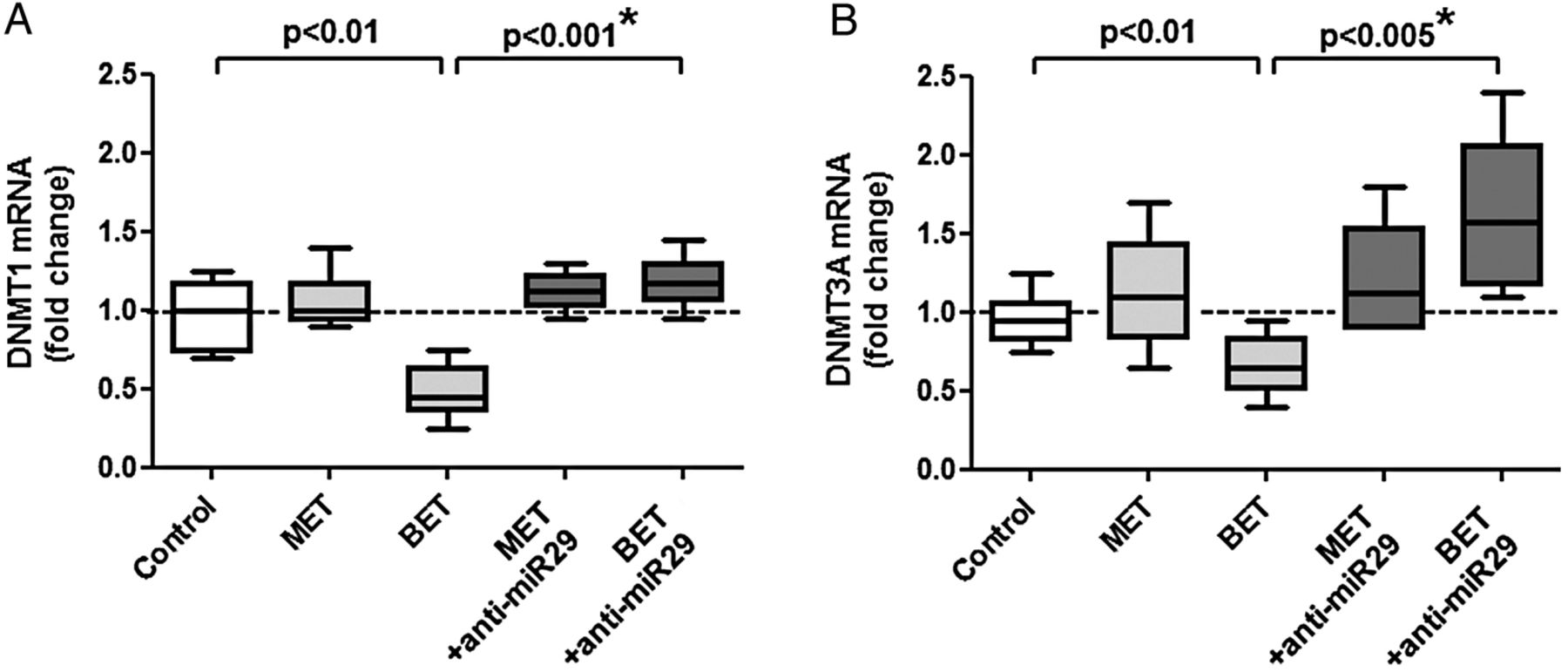
Effects of anti-miR29 on the expression of (A) DNMT1 and (B) DNMT3A transcripts in untreated RASF (control) or treated with L-methionine or betaine. Betaine reduced the expression of DNMT1 and DNMT3A. This can be normalised by treatment with anti-miR29; the significant increase between betaine alone and betaine plus anti-miR29 is confirmed by the analysis of variance. Statistics: Box-and-Whisker plots, Wilcoxon signed rank test between RASF before and after transfection with anti-miR29, *confirmed by Kruskal-Wallis with Dunn's post hoc tests (overall: p<0.005 for (A and B); *confirmed separately for BET vs BET+anti-miR29 regarding DNMT1 and DNMT3A, n=6). BET, betaine; DNMT, de novo methyltransferase; MET, L-methionine; miRs, microRNAs; RASF, rheumatoid arthritis synovial fibroblasts.
Effects of betaine on other cellular processes
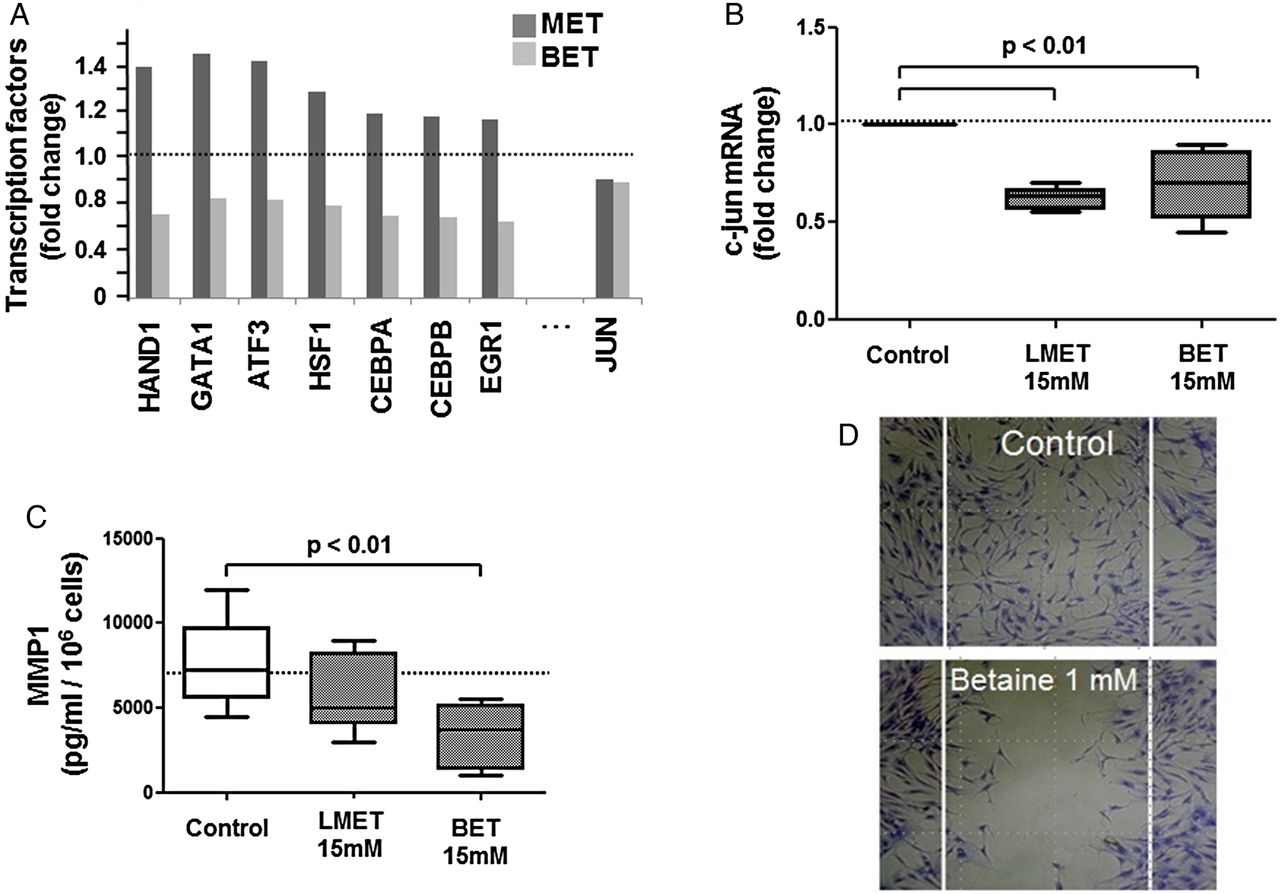
Apparently, betaine was less efficient in remethylating RASF in comparison to L-methionine due to downregulation of DNMT3A by miR29. This is suggested by the observed associations above. On the other hand, betaine has several beneficial effects. Betaine doesn't increase the level of homocysteine (see online supplementary figure S3) and downregulates the expression of multiple transcription factors (figure 6A see online). The c-Jun transcription factor was downregulated by both methyl supplements (figure 6B). Thus, betaine showed multiple beneficial effects unrelated to the DNA remethylation process, which at the end resulted in a modified behaviour of synovial fibroblasts, for example, decreased release of MMP-1 protein (figure 6C) and reduced cell migration (figure 6D).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
(A) The mRNA expression of multiple transcription factors was reduced in betaine-treated RASF, as shown by a transcription factor array; in comparison, L-methionine only reduced c-Jun transcripts. (B) Decreased c-Jun transcript confirmed by real-time PCR. (C) Reduced MMP-1 released by cells treated with L-methionine or betaine. (D) Inhibition of cell migration on a low non-cytostatic dose of betaine. Statistics: bars, Box-and-Whisker plots, Wilcoxon signed rank tests. BET, betaine; MET, L-methionine; MMP-1, matrix metalloproteinase-1; RASF, rheumatoid arthritis synovial fibroblasts.
Discussion
Previously, we showed that an intrinsically enhanced polyamine recycling pathway, controlled by SSAT1, could be responsible for an increased consumption of SAM causing, at long term, the observed DNA hypomethylation in RASF.6 ,7 DNA remethylation of RASF using methyl donors could not be possible in half of the cases without concomitantly inhibiting this pathway.7 However, in some other cases, treatment of RASF with a methyl donor alone was already beneficial, as shown in the SCID mouse model of RA.7 In the current study, we found that miR29 targets DNMT3A and limits DNA remethylation on betaine. Increased SSAT1 levels didn't appear to be responsible for this limitation; however, SSAT1 activity is dependent on the intracellular conditions which can be different in RASF and OASF, as well as before and after treatment with methyl donors; this has to be further investigated. The intracellular competitive/mixed inhibition of SSAT1 activity in RASF could be due to negative feedback product inhibition (ie, increased putrescine and/or diacetylpolyamines). Here, we explored how miRs may selectively target DNA methylation control. Thus, the remethylation efficiency might be limited through upregulation of specific miRs targeting epigenetic effectors.8–12 ,17
In RASF treated with L-methionine, DNMT3B transcripts increased; this could explain at least in part the remethylation. However, the correlation analysis doesn't show a positive relationship. This might be due to a shift in the time course and has to be investigated in more details. Interestingly, in betaine-treated cells, a positive relationship between changes in DNA methylation and DNMT3A transcripts was observed. It might be argued that the change in DNA methylation is low. In RASF, considering that the baseline DNA methylation is about 3.5%, an increase to a ‘normal’ value of 4.2%, as it is achieved with L-methionine, would correspond to a 17–27% increase of methylation of the whole DNA. This is probably the maximum that can be achieved and may primarily concern the remethylation of repetitive sequences.
The levels of DNMTs are often increased in various tumours, possibly playing a role in the hypermethylation of promoter CpG-rich regions of tumour suppressor genes.18 DNMT3A and DNMT3B play also an essential role in early embryogenesis, the stage in which most programmed de novo methylation events occur. They cannot differentiate between unmethylated and hemimethylated CpG sites. Therefore, in differentiated somatic cells, the levels of de novo DNMTs have to be tightly regulated.
In RASF, on betaine, the increase of miR29 correlated with the decrease in DNMT3A transcripts. We validated DNMT3A as a target for miR29 in synovial fibroblasts. The inverse relationship between miR29 and DNMT3A was also described by others in ischaemic brain damages.19 Moreover, miR29 has multiple regulatory functions, for example regulating collagen expression in systemic sclerosis20 and DNMT3A/3B expressions in haematologic malignancies.21 However, miR29 is not upregulated in RA under basal conditions, neither in peripheral blood leucocytes compared with healthy donors,22 nor in synovial fibroblasts compared with OA (data not shown). Nevertheless, under certain conditions, our data suggested that miR29 selectively target mechanisms of DNA methylation.
In spite of the associations reported here, the central question of whether betaine was less efficient than L-methionine, due to downregulation of DNMT3A by miR29, remains partially unanswered. L-methionine and betaine showed similar effects on miR29 expression, the important difference being that in betaine-treated cells DNMT3A is restored to basal levels after transfection with anti-miR29. The question remains how increased miR29 in L-methionine-treated cells doesn't reduce DNMT3A and limit DNA methylation at the same extent? Our results would imply that miR29 only control DNMT3A levels in the presence of betaine. Considering the many effects of betaine unrelated to DNA methylation, in particular on a variety of transcription factors, it might be possible that the effect of betaine is indirect.
The idea to use L-methionine or SAM in the treatment of RA is not novel. In an early clinical study, L-methionine (5 or 10 g/day) was given to patients as a treatment for RA.23 No statistical difference between treatment and controls and no serious adverse effects were reported. Even previously, a double-blinded multicentre study reported that the beneficial effect of SAM is slightly better than ibuprofen in the treatment of hip and knee OA.24 Both drugs were given orally, 0.4 g thrice daily for 30 days. Such observations raise the question of dosage schedule and bioavailability, as well as whether such treatments can reach in vivo the effective and non-cytostatic of dose of 20 mM L-methionine used in vitro. Compared with the free plasma level of L-methionine (about 0.02 mM) or to the interstitial concentration (about 0.11 mM),25 it is 102 to 103 times higher. The intracellular level will depend on the L-methionine transmembrane transporter activity. Higher doses will trigger the conversion into homocysteine. Betaine plasma concentrations increased sharply after oral dosing (50 mg/kg per os), reaching a peak concentration of 0.9 mM after 90 min.26 The clearance is fast, with a T1/2 of about 30 min. Thus, the concentration of 20 mM used in vitro can be considered as very high; however, we already detected changes in cell migration with 1 mM. The local concentrations, however, are certainly higher if a betaine solution is directly injected into the knee joint. Clearly, more information regarding formulation, pharmacokinetic and bioavailability is needed.
The induction of counter-regulatory miRs might represent a general mechanism avoiding an excessive DNA methylation in presence of elevated intracellular betaine levels. It can be hypothesised that repressing the levels of miR29 in RASF during betaine supplementation by using anti-miR29 might enhance the effects of betaine on DNA remethylation.
Betaine inhibited the proliferation of synovial fibroblasts and SW982 sarcoma cells, but not of HepG2 cells, a hepatic tumour cell line that lost the ability to produce BHMT.4 This showed that hepatocytes and synovial fibroblasts express BHMT, allowing a folate-independent methionine metabolism. Betaine, in contrast to L-methionine, decreased homocysteine levels. In RA, this could be beneficial since elevated plasma homocysteine has been associated with increased risk of cardiovascular diseases.27
The c-Jun transcription is downregulated by L-methionine and betaine. Thus, it can be expected that L-methionine and betaine can reduce inflammation and joint destruction in vivo, even independently of DNA remethylation. Betaine downregulates, in addition, mRNAs of multiple transcription factors, such as HAND1, GATA1, ATF3; in particular, the effect on ATF3 transcription could explain betaine's antimigrating and cytostatic properties.28 Clinical trials with betaine could be promising.
References
Footnotes
NG and EK share first authorship.
Contributors Data were acquired by NG, EK, SG and EB. AJ, MF-B and MN analysed the results. Analysis of figures and statistics was performed by MN. BAM accomplished clinical organisation, patient samples, reading and correcting the manuscript. NG, EK and MN wrote the manuscript. AJ, MF-B, REG and SG read and corrected the manuscript. The work was founded by AJ, MN, REG, SG. All authors finally approved the manuscript.
Funding Baugarten Foundation (Zurich, Switzerland).
Competing interests None declared.
Patient consent Obtained.
Ethics approval The procedure was approved by the Ethics Committee of the University Hospital Zurich, and canton of Zurich, Switzerland.
Provenance and peer review Not commissioned; externally peer reviewed.
Data sharing statement Data set will be available from the Dryad repository at http://datadryad.org/