Article Text

Abstract
Objective To determine the association between endoplasmic reticulum aminopeptidase (ERAP)1 and ERAP2 single-nucleotide polymorphisms (SNPs) and human leukocyte antigens (HLA)-B27+ or HLA-B15+ patients with spondyloarthritis (SpA).
Methods 104 patients with SpA according to Assessment of Spondyloarthritis International Society criteria were included in the study. HLA typing was performed by PCR. The polymorphisms were determined by real-time PCR on genomic DNA using customised probes for SNPs rs27044, rs17482078, rs10050860 and rs30187 in ERAP1, and rs2910686, rs2248374 and rs2549782 in ERAP2.
Results 70 of the104 patients with SpA were HLA-B27+ and 34 were HLA-B15+. The distribution of ERAP1 and ERAP2 SNPs between the HLA-B15+ and HLA-B27+ patients with SpA did not reveal differences. Likewise, no differences in the frequencies of ERAP1 SNP haplotypes and alleles HLA-B15 or HLA-B27 were found. Interestingly, however, the frequencies of three particular haplotypes formed by ERAP2 SNPs rs2549782/rs2248374/rs2910686 varied between HLA-B15+ and HLA-B27+ patients: the ERAP2 SNPs haplotype TGT was more common in HLA-B15+ patients with SpA (OR 2.943, 95% CI 1.264 to 6.585; P=0.009), whereas the ERAP2 SNP haplotypes TGC and CAT were more associated with HLA-B27+ patients with SpA: (OR 4.483, 95% CI 1.524 to 13.187; p=0.003) and (OR 9.014, 95% CI 1.181 to 68.807; p=0.009), respectively.
Conclusion An association was found between HLA-B15+ patients with SpA and haplotype TGT of ERAP2 SNPs. On the other hand, HLA-B27+ patients with SpA were associated with ERAP2 haplotypes TGC and CAT. These associations could be related to the clinical presentation of the disease, specifically with a peripheral or axial predominance, respectively.
- Spondyloarthritis
- Gene Polymorphism
- Ankylosing Spondylitis
- Autoimmune Diseases
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
INTRODUCTION
Spondyloarthritis (SpA) are a heterogeneous group of chronic inflammatory joint diseases, characterised by axial and peripheral manifestations, as recognised by the implementation of the new classification criteria proposed by the group Assessment of Spondyloarthritis International Society (ASAS). In the axial presentation, the sacroiliac joints are particularly affected, followed by an ascending compromise of the spine. The peripheral manifestations are characterised by asymmetric oligoarthritis and enthesitis of the lower limbs.1–3
The origin of SpA is considered to be multifactorial, where a genetic susceptibility background coupled with the presence of certain environmental triggers stimulates the onset of the disease.4 Similar to other rheumatic diseases, a polygenic origin has been proposed, where the strongest association is established with molecules of the major histocompatibility complex (MHC-I), specifically the human leukocyte antigen B27 (HLA-B27).5 While HLA-B27 molecule is recognised to confer genetic susceptibility to develop SpA (with positivity in about 25–40% of patients), and several hypotheses have been proposed to explain its participation in the development of the disease, none of them clearly explain the role of this molecule for the pathogenesis of the disease.5
Advances in genetics and studies in large populations have additionally established associations of more than 40 genes unrelated to the MHC and the development of SpA. The most consistent of them are the genes of the endoplasmic reticulum aminopeptidase (ERAP) type 1 and ERAP2.6 7 These enzymes include a single class of pleiotropic proteases with localisation in the endoplasmic reticulum (ER). They play an important role in the process of N-terminal fraction of the antigenic peptide that will occupy the MHC-I molecules for presentation at the cell surface to CD8+ lymphocytes.8–11
ERAP1 and ERAP2 have unique peptidic specificities. Evidence indicates that ERAP2 is involved in antigenic cleavage in ER and forms a repertoire of ligands for MHC-I molecules that are different from those generated by ERAP1. This leads to an efficient and coordinated presentation of a wide variety of peptides when these enzymes act together.12
The association of ERAP1 polymorphisms with SpA appeared to be related with the predisposition to develop the disease in HLA-B27+ individuals and ERAP2 polymorphisms in individuals with other non-HLA-B27 alleles. However, Robinson et al found an association of ERAP2 polymorphisms with ankylosing spondylitis (AS) in both HLA-B27+ and HLA-B27− patients.13
Currently, our research group is focused on the particular genetics and phenotypes of the patients with SpA of Latin America ancestry (in Colombia). Previously, we analysed the presence of HLA-A, HLA-B and HLA-DR molecules in 189 Colombian patients with a diagnosis of SpA plus 100 healthy subjects who were on the list of transplant donors, and had similar socio-demographic characteristics as our SpA population. In addition to the expected increase in the frequency of HLA-B27, we also found an increased frequency of HLA-B15 in our patients with SpA.14 Also, we reported a clear association between HLA-B27 with axial SpA including AS, and HLA-B15 with peripheral SpA including undifferentiated spondyloarthritis.15
The objective of the present study was to establish the association of polymorphisms of ERAP1 and ERAP2 genes in Colombian patients with SpA with HLA-B27+ or HLA-B15+, and to evaluate how this association could be related to the clinical presentation of the disease, specifically with axial or peripheral predominance.
MATERIALS AND METHODS
Patients and clinical assessment
A group of 104 patients with diagnosis of SpA according to the European Spondyloarthropathy Study Group (ESSG) criteria16 and positive for either HLA-B27 or HLA-B15 were included in the current study of ERAP SNPs. Patients were collected from an outpatient clinic of SpA of two referral hospitals in Bogotá, from January 2009 to December 2016. Subsequently, patients were classified according to the new ASAS criteria for the classification of the axial and peripheral SpA. Demographic and clinical data, including Bath Ankylosing Spondylitis Disease Activity Index (BASDAI),17 functional capacity (Bath Ankylosing Spondylitis Functional Index (BASFI)),18 global disease activity assessed by physicians and by patients through a visual analogue scale (VAS), and joints affected were recorded.10 Mander’s index was used to evaluate involved enthesis inflammations.19 For each patient, routine laboratory tests were assessed.
In a previous study,14 our patients with SpA and a control group of 100 healthy subjects on the list of transplant donors with similar socio-demographic characteristics as our SpA population were compared for the analysis of the Hardy-Weinberg equilibrium by using the Arlequin statistical software version 3.5.20 Allele frequency (AF) among the patient population was calculated using updated versions of the Genepop Program.21 Likewise, the increase in the frequency of HLA-B27 or HLA-B15 in the group of patients with SpA, compared to the aforementioned healthy control population, was determined with the Genepop Program as described.14
Imaging
In all patients, a plain anterior pelvic X-ray and MRI of the sacroiliac joints with T1 sequence and fat suppression technique were obtained. The images were evaluated by a radiologist with musculoskeletal expertise who was blinded to the clinical information. The X-ray readings were made following the recommendations for population studies according to the Atlas of Standard Radiographs.22 MRI was evaluated looking for acute changes in the joints.23
HLA genotyping
Genomic DNA was extracted from peripheral blood leukocytes with a Wizard Genomic DNA Purification Kit (Promega, Cat. # A1120). After extraction, samples were adjusted to a DNA concentration of 80±20 μg/μL and stored at −80°C until processing of HLA. Typing was carried out by PCR on the genomic DNA using the HLA Sequence-Specific Primer HLA-ABDR-SSP tray Kit (BioRad, Cat. # 826 230), which is focused on determining HLA-A, B, DRβ1 alleles. PCR products were verified by electrophoresis in a 2% agarose gel, and bands were visualised by ultraviolet light photo documentation and further analysed by using the HLA-SSP Typing V 1.2.0.0. Software (BioRad).
Inflammatory serological biomarkers
A series of inflammatory serological biomarkers could be measured in a subset of patients as follows: the levels of tumour necrosis factor α, interleukin (IL)-1α, IL-6, and IL-17 were measured by a cytometric bead-array (CBA Flex Set) using a FACSCanto II Flow Cytometer. ELISA was used to determine the serum levels of IL-23, macrophage colony-stimulating factor and metalloproteinase 3. C reactive protein and lipopolysaccharide-binding protein levels were measured by chemiluminescence.
ERAP polymorphisms
Identification of single-nucleotide polymorphisms (SNPs) was performed by the technique of real-time PCR. TaqMan-labelled primers of ERAP1 (rs27044, rs30187, rs10050860, rs17482078) and ERAP2 (rs2549782, rs2248374 and rs2910686) SNPs were designed and synthesised by Roche. The TaqMan genotyping method was then applied to analyse all SNPs using the LightCycler 480 ROCHE equipment, and the results were analysed by LightCycler 480 software. The characteristics and references of the analysed SNPs of ERAP1 and ERAP2 are shown in table 1.
Polymorphisms of ERAP1 and ERAP2 genes in HLA-B27+ and HLA-B15+ patients with SpA
In a two-step exercise, each SNP was initially tested in the two groups of HLA-B27+ and HLA-B15+ patients with SpA and then, based on the principle of non-independent gene segregation, the potential haplotypes formed by the ERAP1 and ERAP2 SNPs were analysed in relation to HLA genes. To determine haplotype frequencies for ERAP SNPs in HLA-B7+ and HLB-B15+ patients, the Haploview software was used.24
Statistical analysis
Arlequin,20 Genepop and Haploview24 softwares were used as mentioned earlier. For analysis of data, we used the statistical package SPSS 19.0 (SPSS, USA). For the presentation and analysis of categorical variables, frequency distribution, percentages and contingency tables were used. Comparisons were made by the χ2 test and the Fisher’s exact test. For numerical values, central tendency and dispersion measurements were evaluated. Results were analysed using Student’s t-test and Wilcoxon test. Two-tailed p values <0.05 were considered statistically significant. The association between disease and HLA alleles was determined by OR and 95% CIs.
RESULTS
A total of 70 patients with SpA with HLA-B27 and 34 patients with SpA with HLA-B15 were genotyped for SNPs of ERAP1 and ERAP2 genes. A male predominance (male to female ratio of 3.7:1) was documented in HLA-B27+ group, while in HLA-B15+ patients, the gender ratio was 0.8:1. The demographic and clinical information is shown in table 2. Based on the ASAS classification criteria, 97 patients met the criteria for axial SpA, and 94 patients fulfilled the ASAS criteria for peripheral SpA (see table 2). Notably, 100% of the 70 HLA-B27+ patients with SpA included in the study exhibited axial manifestations of the disease and 100% of the 34 HLA-B15+ patients with SpA had peripheral manifestations.
Characterisation of demographic and clinical variables in patients with SpA with HLA-B27 and HLA-B15
As previously reported,15 patients with HLA-B27 displayed higher sacroiliitis grade II when compared to HLA-B15+ patients with SpA (74.7% vs 56.3%, p=0.043). Also, higher enthesitis in the sacroiliac region was found in 64.4% of HLA-B27+ patients compared to 36% of HLA-B15+ patients (p=0.001). On the other hand, patients with HLA-B15 had higher number of swollen joints (5.3±5.1 vs 2.3±1.8; p=0.004), higher count of painful enthesitis (6.0±4.5 vs 4.3±3.3; p=0.014) and painful peripheral enthesitis (3.4±2.9 vs 2.3±2.2; p=0.050) than HLA-B27+ patients. Moreover, HLA-B15+ patients had higher scores on BASDAI and BASFI, and overall disease activity assessed by the physician (VAS physician) when compared to HLA-B27+ patients, while HLA-B27+ patients had a higher VAS score in dorsal inflammatory pain and VAS of overall disease activity assessed by the patient.
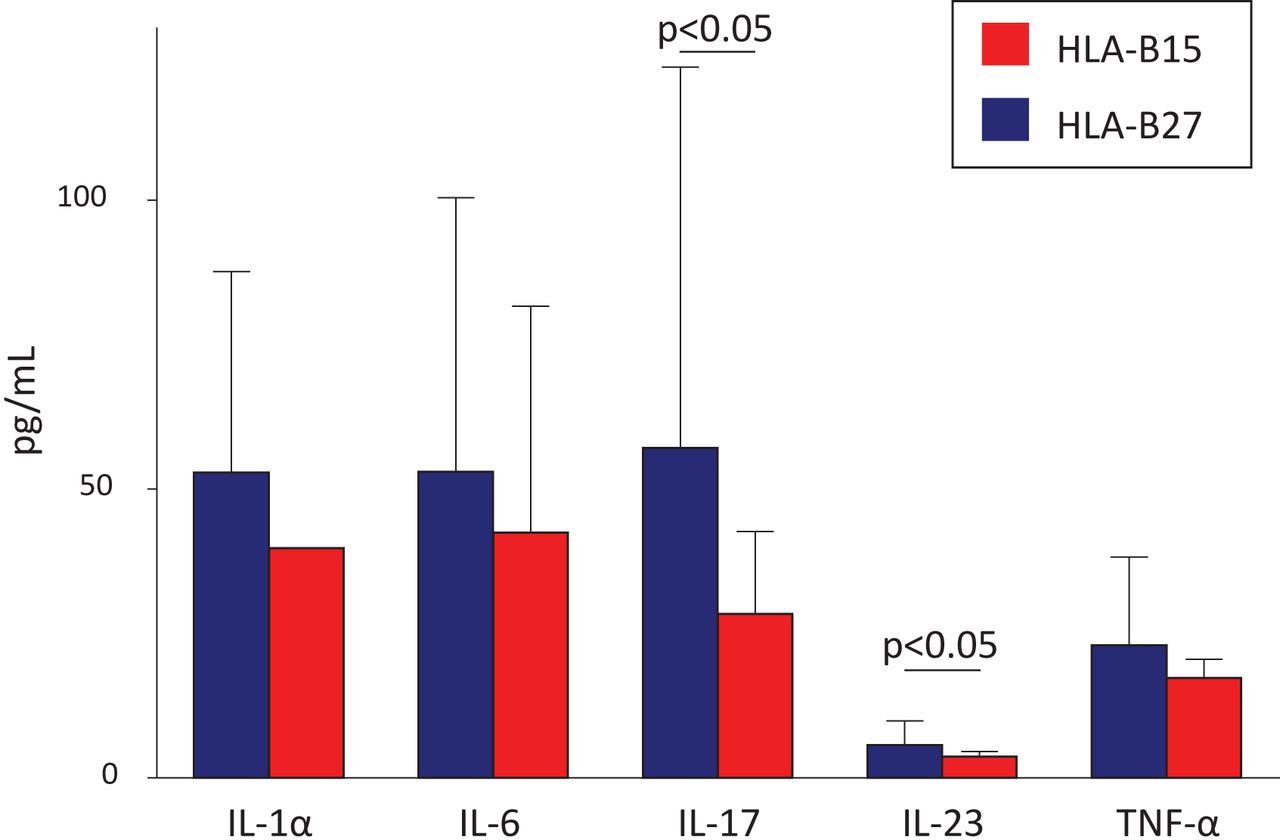
These aforementioned clinical and radiological differences previously encountered in SpA between HLA-B27+ and HLA-B15+ patients may correlate with our current preliminary findings related to serological biomarkers we managed to obtain in a small subset of patients: for most biomarkers, a trend to higher levels in the HLA-B27+ subset of patients versus the HLA-B15+ subset was observed (table 3). Interestingly, and despite the relatively low number of patients analysed, this trend turned into statistically significant difference in the case of two biomarkers compromised with immunopathogenesis of SpA: IL-23 (5.7±4.2 vs 3.7±0.9 pg/mL; p<0.05) and IL-17 (57.1±66.0 vs 28.4±14.3 pg/mL; p<0.05) (figure 1).
Levels of inflammatory serological biomarkers in patients with SpA according to the presence of HLA-B27 or HLA-B15
{kind=link}
Levels of serum cytokines in patients with SpA positive for HLA-B27 (n=25) or HLA-B15 (n=12). Results are expressed as mean±SD. HLA, human leukocyte antigen; IL, interleukin; SpA, spondyloarthritis; TNF-α, tumour necrosis factor α.
The distribution of each SNP of the ERAP1 and ERAP2 genes between the HLA-B15+ and HLA-B27+ patients did not reveal any significant difference in AFs (not shown). However, when we analysed the frequencies of all possible haplotypes formed by the respective ERAP SNPs and the alleles HLA-B15 or HLA-B27, important associations were observed regarding three haplotypes of the ERAP2 SNPs rs2549782/rs2248374/rs2910686 (table 4). Thus, the haplotype TGT was found to be more frequent in HLA-B15+ patients with SpA, and two additional haplotypes (TGC and CAT) were more frequent in HLA-B27+ patients with SpA. On the other hand, we did not observe differences in the frequency of haplotypes generated from the ERAP1 SNPs and alleles HLA-B15 or HLA-B27 (table 4), suggesting that SNPs of ERAP1 are not associated with a disease-specific phenotype in our Colombian patients with SpA.
Frequency of ERAP SNP haplotypes in patients with SpA with HLA-B27 and HLA-B15
DISCUSSION
Based on the presumed polygenic origin of SpA, we analysed the differences in the frequencies of the haplotypes formed by ERAP1 and ERAP2 SNPs in relation to the presence of HLA-B27 or HLA-B15 alleles in our Colombian patients with SpA. Interestingly, we found differences in the frequencies of three ERAP2 SNP haplotypes, one of them associated with positivity for HLA-B15 and two with HLAB27 positivity.
In 2007, a wide association scan of multiple nonsynonymous SNPs identified association of three different SNPs of ERAP1 with AS.6 Shortly afterwards, Maksymowych et al studied 992 Canadian patients with AS and healthy unrelated controls, most of them Caucasians of northern European descent: a specific ERAP1 haplotype, rs27044/rs10050860/rs30187 CCT, was found to be associated with increased risk of AS (OR 1.81, 95% CI 1.46 to 2.24; p=0.007×10−5), while a second specific ERAP1 haplotype, rs30187/rs26618/rs26653 CTG, reduced the disease risk (OR 0.77, 95% CI 0.67 to 0.88; p=0.009×10−2).25 The observed ERAP1-AS association was therefore initially limited to patients with SpA and HLA-B27+, and indeed it appeared to support the polygenic origin of the disease and would have important implications.6 26 The involvement of ERAP1 and HLA-B27, two molecules intimately linked in the processing and antigenic presentation, could affect the stability of HLA–antigen complex and modulate the repertoire of antigenic peptides, presented to the immune system. This interesting association could not be applied to patients with SpA negative for HLA-B27, a frequent situation in non-Caucasian populations.
In 2013, the results of the International Genetics of Ankylosing Spondylitis Consortium (IGAS) study were published. This study involved 10 619 individuals with AS and 15 145 controls of European, East Asian and Latin American ancestry. The researchers found association of AS with two SNPs in ERAP2, rs2549782 and rs2248374, in HLA-B27+ patients. After controlling the ERAP1 interaction over ERAP2 polymorphism, they observed association with ERAP2 SNPs in HLA-B27− AS cases (rs2910686: OR 1.19, p=0.213×10−2).27
The study of Costantino et al in 2015 ratified the previously reported association of ERAP1 haplotype (rs17482078, rs10050860, rs30187) and the susceptibility of developing AS.28 More recently, Babaie et al studied 220 Iranian patients with AS and unrelated controls, and confirmed that two ERAP1 SNPs (rs27044 and rs10050860) are associated with AS also in this population.29 On the other hand, Su et al did not find any association between some ERAP1/ERAP2 SNPs and AS in Chinese patients.30
In line with these later results, we did not find independent associations of ERAP1 or ERAP2 SNPs with SpA in our HLA-B15+ or HLA-B27+ patients (not shown). Interestingly, however, we observed different frequencies of three distinct haplotypes of the analysed ERAP2 SNPs (rs2549782, rs2248374, rs2910686) between HLAB15+ and HLAB27+ patients with SpA. The haplotype TGT of the ERAP2 SNPs was more frequent in HLA-B15+ patients, while two additional haplotypes of the same ERAP2 SNPs (TGG and CAT) were more frequent in HLA-B27+ patients with SpA (table 4).
Previously, missense SNPs in ERAP2 (one of them included in our current study) have been linked with predisposition to autoimmune diseases. It has been hypothesised that altered antigen processing by this enzyme is a causal link to disease aetiology. Evnouchidou et al reported in 2012 that the nonsynonymous ERAP2 SNP, rs2549782—T/G (the T allele coding for 392 N, and the G allele codes for 392 K variant of ERAP2; cf. Table 1) leads to alterations in both the activity and the specificity of the enzyme. Particularly, the 392 N variant excises hydrophobic N-terminal residues from epitope precursors up to 165-fold faster compared with the 392 K variant, while both variants are very similar in excising positively charged N-terminal amino acids. X-ray crystallographic analysis suggests that the polymorphism interferes with the stabilisation of the N-terminus of the peptide through interactions with key residues participating in the catalytic activity of the enzyme. The change leading to a more active enzyme variant may allow ERAP2 to supplement ERAP1 activity for the removal of hydrophobic N-terminal residues. On the other hand, it may be noticed that rs2549782 is in linkage disequilibrium with rs2248374 and that the change from A to G allele is deleterious for ERAP2 mRNA splicing, leading to mRNA decay, and deficient expression of the enzyme.31 Altogether, these data provide mechanistic insights to the association of ERAP2 polymorphisms with the disease and support the idea that polymorphic variation in antigen-processing enzymes constitutes a component of the immune response variability in humans.32
Variations in the length and chemical composition of the antigenic peptides and the subsequent binding of aberrant peptides to the MHC-I molecules may result in the formation of unstable bonds between HLA molecules with the antigenic peptide.33 Ultimately, it would lead to accumulation of ‘misfolded MHC-I’ molecules in the ER, inducing stress and generating a pro-inflammatory cellular response called unfolded protein response.34 35 These alterations might condition the intensity of the immune response, which in the case of SpA, would be in relation to the clinical characteristics of the disease, with predominance of the axial symptoms or with predominance of the peripheral symptoms.
The possibility of modulating the inflammatory response because of interaction of ERAP2 SNPs with HLA-B27 and HLA-B15 genes in patients with SpA is interesting. The polygenic action of ERAP-HLA can increase or decrease the grade of response and modulate the clinical presentation of the disease. Our current preliminary data of a significant increase in the levels of IL-23 and IL-17 in patients with SpA with HLA-B27 versus those with HLA-B15 (figure 1 and table 3) may provide useful information on the pathogenic relevance of the aforementioned comments.
The results of previous research have confirmed that the presence of the HLA-B27 allele is related to the development of a predominantly axial disease, whereas the HLA-B15 allele is associated with predominantly peripheral disease.14 This situation offers a clinical model that can help to establish if the genetic susceptibility influences the SpA phenotype.36–38 The results obtained in the present investigation seem to support this hypothesis.
Further studies should be carried out to establish the true interaction of genes involved in the susceptibility of developing disease and the modulation of the chronic inflammatory response. The results obtained could favour the development of selective therapeutic targets in patients with SpA according to compromised autoimmune mechanisms.
Key messages
What is already known about this subject?
ERAP1 and ERAP2 polymorphisms have been associated with SpA: ERAP1 SNPs preferentially with HLA-B27+ patients and ERAP2 SNPs with HLA-B27− patients.
What does this study add?
In this study, conducted on a group of Latin American patients with SpA, particular ERAP2 but not ERAP1 SNP haplotypes were found to be associated with either HLA-B27+ or HLA-B15+ patients with SpA.
The results of previous research have confirmed that the presence of the HLA-B27 allele is related to the development of a predominantly axial disease, whereas the HLA-B15 allele is associated with a predominantly peripheral disease.
How might this impact on clinical practice?
The association of ERAP2 SNP haplotypes with HLA-B27+ or with HLA-B15+ patients with SpA could condition the pattern of clinical presentation of the disease, preferentially axial or peripheral disease.
The results of this study deserve to be validated further with other studies involving a larger number of patients with SpA.
REFERENCES
Footnotes
Twitter Santiago Bernal-Macías @BernalMacias90.
Acknowledgements We are grateful to Maria del Pilar Delgado, MCs, Department of Biological Sciences, University of Los Andes, for critical review of the manuscript.
Contributors All authors were involved in drafting the article or revising it critically for important intellectual content, and all authors approved the final version to be published. JL had full access to all the data in the study and takes responsibility for the integrity of the data and the accuracy of the data analysis. Study conception and design: JL, AMS, JCR, EC-P, RB-V, GV-A, NM-R, FC, LMA-P, CR, JFM. Acquisition of data: JL, AMS, JCR, EC-P, SA-C, G-NM, DP, FC, VR-M, SB-M, CV-E. Analysis and interpretation of data: JL, AMS, JCR, EC-P, RB-V, GV-A, NM-R, SA-C, G-NM, DP, VR-M, SB-M, CV-E, LMA-P, CR, JFM.
Funding This work was supported by the University of La Sabana (Med-232-2018) and Asociación Colombiana de Reumatología (ASOREUMA) (Acta 019/2014).
Disclaimer These sponsors had no role in study design, data analysis or the decision to publish.
Competing interests None declared.
Ethics approval The study followed the norms established by the Helsinki Declaration, The Guidelines for Good Clinical Practice, and the Resolution 8430 (1993) of the Colombian Ministry for Social Protection. Moreover, the study was reviewed and approved by the Ethics Committees of the University of La Sabana, Los Andes University and the Central Military Hospital. Each patient signed an informed consent form, and confidentiality was strictly maintained.
Provenance and peer review Not commissioned; externally peer reviewed.
Data availability statement Data are available on reasonable request. All data relevant to the study are included in the article.