Article Text

Abstract
Background Perifascicular atrophy is a unique pathological hallmark in dermatomyositis (DM)-affected muscles; however, the mechanism underlying this process remains unclear. In this study, we aimed to investigate the potential role of the immunoproteasome subunit β5i and retinoic acid-inducible gene-I (RIG-I) in DM-associated muscle atrophy.
Methods The expression of β5i and RIG-I in the muscles of 16 patients with DM was examined by PCR, western blotting and immunohistochemistry. The associations between β5i and RIG-I expression levels and muscle disease severity were evaluated. Lentivirus transduction was used to overexpress β5i in human skeletal muscle myoblasts (HSMMs) and consequent cell functional changes were studied in vitro.
Results β5i and RIG-I expression in the muscle of patients with DM was significantly increased and closely associated with muscle disease severity. Immunohistochemistry and immunofluorescence analyses showed the marked colocalised expression of β5i and RIG-I in perifascicular myofibres. β5i overexpression in HSMMs significantly upregulated RIG-I, the muscle atrophy marker MuRF1, type I IFN-related proteins (MxA and IFNβ) and NF-κB pathway-related proteins (pIκBα, pIRF3 and pNF-κBp65). In addition, the viability of HSMMs decreased significantly after β5i overexpression and was partly recovered by treatment with a β5i inhibitor (PR957). Moreover, activation of RIG-I by pppRNA upregulated IFNβ and MuRF1 and reduced the cell viability of HSMMs.
Conclusion The immunoproteasome subunit β5i promotes perifascicular muscle atrophy in DM via RIG-I upregulation; our findings suggest a pathomechanistic role of β5i and RIG-I in DM-associated muscle damage, highlighting these components as potential therapeutic targets for the treatment of DM.
- Dermatomyositis
- Autoimmune Diseases
- Immune System Diseases
Data availability statement
Data are available on reasonable request.
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
WHAT IS ALREADY KNOWN ON THIS TOPIC
Perifascicular atrophy is a pathological hallmark of dermatomyositis (DM)-affected muscles.
WHAT THIS STUDY ADDS
The immunoproteasome subunit β5i promotes perifascicular muscle atrophy in DM via RIG-I upregulation.
HOW THIS STUDY MIGHT AFFECT RESEARCH, PRACTICE OR POLICY
The pathomechanistic role of β5i and RIG-I provides insights into the mechanisms underlying DM-associated muscle damage and underscore a new potential therapeutic target for DM.
Introduction
Dermatomyositis (DM), a disease involving multiple organ systems, is characterised by skeletal muscle inflammatory damage. Perifascicular atrophy is a pathological hallmark of DM-affected muscles; however, the mechanisms underlying its occurrence and development remain unclear.1
The marked overproduction of type I interferon (IFN)-inducible transcripts and proteins, which are preferentially located in perifascicular myofibres, indicates that type I IFN is closely related to perifascicular atrophy.2 3 Retinoic acid-inducible gene-I (RIG-I) is a receptor associated with innate immunity; RIG-I recognises and binds to double-stranded viral RNA molecules, forming a positive feedback loop associated with type I IFN.4–6 We previously reported that RIG-I expression increased significantly in peripheral T lymphocytes and participated in the mechanisms underlying T cell lymphopenia in patients with DM.7 Meanwhile, a study showed that RIG-I is overexpressed in perifascicular myofibres in DM.8 Nonetheless, whether aberrantly expressed RIG-I plays a pathogenic role in DM-associated perifascicular damage remains unclear.
Immunoproteasome plays a critical role in processing antigens for their presentation on major histocompatibility complex-I (MHC-I), regulating the production of proinflammatory cytokines and maintaining protein homoeostasis.9 A previous study showed that elevated levels of the immunoproteasome subunit β5i in muscle tissues were involved in pathological MHC class I expression and the maintenance of myokine production in DM.10 In addition, several reports have suggested that the immunoproteasome is largely responsible for inflammation-induced muscle proteolysis and aged muscle atrophy.11–13 Therefore, whether β5i participates in DM-associated muscle atrophy is worthy of investigation.
In this study, we comprehensively investigated the expression of β5i and RIG-I in DM-affected muscle tissues and analysed their relationship with the severity of skeletal muscle disease. Furthermore, we explored the potential roles of β5i and RIG-I in myoblast atrophy in vitro.
Materials and methods
Patients and controls
Sixteen hospitalised treatment-naive patients with DM admitted to the Department of Rheumatology in the China Japan Friendship Hospital between March 2019 and January 2020 were enrolled in this study. The classification criteria for DM were based on the 2017 EULAR/American College of Rheumatology Classification Criteria for Adult Idiopathic Inflammatory Myopathies.14 These 16 patients did not show complications, such as overlap syndrome, infectious diseases and cancer. Controls of skeletal muscle tissues were obtained from five orthopaedic trauma patients without a medical history of skeletal diseases.
Clinical characteristics
Clinical characteristics, including sex, age, rash (heliotrope rash, Gottron rash, periungual erythema and ‘mechanic’s hand’), interstitial lung disease (ILD) and oesophageal lesions, were assessed. The muscle disease activity was assessed using a continuous 10 cm Visual Analogue Scale (VAS).15 Serum creatine kinase (CK) levels were included for analysis at the clinic visit closest to the time of muscle pathology evaluation. Muscle strength was measured using the manual muscle test (MMT8) proposed by the International Myositis Outcome Assessment Collaborative Study (http://www.niehs.nih.gov/research/resources/imacs/diseaseactivity/index.cfm). The definitions of ILD were in accordance with the policies of the 2013 American Thoracic Society and European Respiratory Society.16 Dysphagia, nasal or gastro-oesophageal regurgitation and aspiration pneumonia were identified as oesophageal lesions; further examinations, including oesophageal manometry, barium-swallow examination or endoscopic examination, were performed to evaluate the oesophageal function, if necessary. Oesophageal lesions were defined according to clinical symptoms and auxiliary examination. Myositis-specific antibodies (MSAs) and myositis-associated antibodies (MAAs), which were detected by immunoblotting analyses using a diagnostic kit (EUROIMMUN, Lübeck, Germany), were assessed. The MSAs and MAAs analysed included anti-Mi-2, anti-TIF1γ, anti-MDA5, anti-NXP2, anti-SAE1, anti-HMGCR, anti-SRP, anti-synthetase (anti-Jo-1, anti-PL-7, anti-PL12, anti-EJ and anti-OJ), anti-Ku, anti-Ro52, anti-PM-Scl100 and anti-PM-Scl75 antibodies.
Muscle pathology evaluation
Assessment of histological severity was performed with reference to a juvenile DM biopsy score tool, which was proposed by Hemlata17 18 with some modifications (online supplemental table 1). This scoring system is a semiquantitative assessment of histological features comprising four domains (muscle fibre, inflammatory, vascular and connective tissue). The muscle fibre domain consists of perifascicular atrophy, MHC class I overexpression and necrosis. As T cells and B cells are predominant lymphocytes involved in the immunopathology of adult DM, the inflammatory domain includes CD3+ and CD20+ endomysial/perimysial infiltration without CD68+ macrophages. Based on the presence or absence of corresponding lesions, the connective tissue and vascular domains were scored as 1 or 0, respectively. The muscle pathological score was obtained by summing each domain score. The scoring process was performed by an experienced pathologist and a rheumatologist blinded to other clinical information. The mean of scores from both specialists was taken as the final score.
Supplemental material
RNA isolation and real-time PCR analysis
RNA was extracted using a TRIzol kit (Thermo, Carlsbad, California, USA), following the manufacturer’s instructions. The RNA samples were reverse-transcribed using the TransScript First-Strand cDNA Synthesis SuperMix kit (Takara, Dalian, Liaoning, China). Quantitative real-time reverse transcriptase PCR was performed using the SYBR Green kit (Takara, Dalian, China) on an ABI 7500 system (Applied Biosystems, Singapore). The PCR analysis was performed according to the manufacturer’s instructions. Each sample was analysed in triplicate. The relative mRNA expression levels of each gene of interest were normalised to the mRNA levels of the housekeeping gene GAPDH. The gene expression levels were calculated according to the 2−ΔCT method. The primer sequences for the genes were as follows: human β5i, forward: 5′-CTGGGTCCTACATTAGTGCCT-3′ and reverse: 5′-TTCTCCATTTCGCAGATAGTACA-3′; human RIG-I, forward: 5′-TGAGTAGACCACATCCCAAGC-3′ and reverse: 5′-GCAATATCCTCCACCACAAAA-3′; GAPDH, forward: 5′-GAGAAGGCTGGGGCTCATTTGCA-3′ and reverse: 5′-TTGGCCAGGGGTGCTAAGCAGT-3′. The primers were synthesised by Invitrogen (USA).
Western blotting analysis
The total proteins were extracted and separated by 10% or 15% SDS-PAGE and were transferred to polyvinylidene difluoride membranes (Millipore, Bedford, Massachusetts, USA) using a semi-dry Gel Transfer Device (Bio-Rad, Hercules, California, USA). The membranes were blocked using 5% non-fat milk and probed with primary antibodies and HRP-conjugated secondary antibodies. Antigen-antibody complexes were visualised using a chemiluminescent ECL detection system and analysed using a ChemDoc XRS+image analyser. GAPDH or β-actin was used as an internal control. The antibodies used included anti-β5i (1:5000, ab180606, Abcam), anti-RIG-I (1:2000,3743, Cell Signaling Technology), anti-Phospho-NF-κB p65 (1:1000, 3033, Cell Signaling Technology), anti-Phospho-IRF3 (1:500, ab76493, Abcam), anti- IFNβ (1:500, ab275880, Abcam), anti- MxA (1:500, sc-166412, Santa Cruz), anti-MuRF1 (1:2000, ab172479, Abcam), anti-β-actin (1:5000, YM3028, Immunoway) and anti-GAPDH (1:10000, YM3029, Immunoway). The selective β5i inhibitor PR-957 was purchased from Selleck Chemicals (Houston, Texas, USA).
Immunohistochemistry
Frozen muscle sections were incubated with H2O2 (3%) for 15 min, followed by washing three times with phosphate buffer solution Tween-20 (PBST). The slides were blocked with goat serum at room temperature for 2 hours. Rabbit antibodies against human β5i (1:2000, ab180606, Abcam) and RIG-I (1:500, ab45428, Abcam) were used as the primary antibodies. A monoclonal rabbit IgG (1:2000, ab37415, Abcam) was used as isotype control for β5i (1:2000, ab37415, Abcam) and a polyclonal rabbit IgG (1:500, ab172730, Abcam) was used as isotype control for RIG-I. After washing with phosphate-buffered saline (PBS), the slides were incubated with HRP-conjugated secondary antibodies. Each incubation step was followed by three washes with PBST. Diaminobenzidine was used as the chromogen to visualise the proteins.
Immunofluorescence
To explore the colocalisation of β5i and RIG-I, double-label immunostaining was performed. After blocking with 5% bovine serum albumin in PBST at room temperature for 1 hour, the samples were treated with mouse anti-RIG-I monoclonal antibody (1:50, sc-376845, Santa Cruz Biotechnology) and rabbit anti-β5i monoclonal antibody (1:2000, ab180606, Abcam) overnight at 4°C, followed by three washes with PBS (10 min each wash). Next, the samples were treated with Alexa Fluor 594-conjugated goat anti-mouse IgG (1:200, 8890, Cell Signaling Technology) and Alexa Fluor 488-conjugated goat anti-rabbit IgG (1:200, 4412, Cell Signaling Technology), which were used as the secondary antibodies, in a dark chamber for 1 hour, followed by three washes with PBST. After counterstaining with DAPI (Beyotime, Shanghai, China), the slides were mounted with coverslips. To clarify whether β5i is expressed in regenerative myocytes, we performed double-label immunostaining using rabbit anti-β5i monoclonal antibody (1:2000, ab180606, Abcam) and mouse anti-NCAM1 monoclonal antibody (1:500, ab6123, Abcam), following the same procedure mentioned above. In addition, double-label immunofluorescence staining using rabbit anti-β5i monoclonal antibody and mouse anti-MxA monoclonal antibody (1:100, sc-166412, Santa Cruz) was performed following the above procedure.
Plasmid construction, lentivirus packaging and transduction
The β5i overexpression and control lentivirus plasmids were purchased from VectorBuilder (Guangzhou, Guangdong, China). The vectors encoded both a red fluorescent protein and a puromycin-resistant protein. The packaging vectors (pLP1, pLP2 and pLP/VSVG, kept by our laboratory) were prepared as described in a previous study.19 To obtain the lentiviruses, recombinant β5i plasmids were cotransfected with the packaging vectors into 293 T cells using Lipofectamine 2000 (Life Science, Carlsbad, California, USA). The supernatants (containing the virus particles) were harvested at 48 hours and 72 hours after transfection. The viruses were then concentrated by loading the samples onto columns (Millipore, Bedford, Massachusetts, USA). The titres of the virus suspensions were determined by serial dilution.
For lentivirus transduction, human skeletal muscle myoblasts (HSMMs; Lonza Bioscience, Basel, Switzerland) were seeded into a six-well plate (1×105 cells/well). The next day, cells were incubated with 5–10 µL of the virus solution diluted in 1 mL of cell culture medium at a multiplicity of infection of 20. Three days later, the transduction efficiency of the virus was evaluated by fluorescence microscopy. Cells showing stable β5i overexpression were established by subjecting them to selection pressure using puromycin (2 mg/mL) for 1 week.
Cell treatments
For PR957 treatment, HSMMs were seeded into six-well plates (Nunc, Roskilde, Denmark). Next, PR957 (Selleck, Houston, Texas, USA), at a concentration of 100 nM was added to the culture medium.
pppRNA stimulation
The cells were transfected with an RIG-I-specific ligand pppRNA (Invitrogen, San Diego, USA) at a concentration of 0.5 µg/mL using the Lipofectamine 3000 transfection reagent (Thermo, USA).
Cell viability assay
The viability of HSMMs was measured using a CCK-8 kit (Dojindo Laboratories, Japan) according to the manufacturer’s instructions. The optical density (OD) values at 450 nm were measured by microplate reader and the results were averaged from five duplicate wells of each group.
Statistical analysis
Statistical analyses were performed using PASW statistics V.18 (IBM). Data are expressed as means±SDs. Comparisons between groups were performed using the independent-samples t-test. Correlation analysis for the variables of interest was performed using Pearson correlation or Spearman rank correlation test when appropriate. A two-sided p value 0.05 was considered to indicate statistical significance.
Results
Baseline characteristics of patients with DM
The age of the patients ranged from 18 to 71 years, with an average of 46.2 (±14.2) years. The average age of healthy control (HC) was 50.4 (±12.7) which was not significantly different from that of patient with DM. The average disease duration was 4.0 months (±2.25, range 2–8 months). These subjects included four patients who tested positive for anti MDA5, two patients for anti-NXP2, one patient for anti-Jo-1, two patients for anti-SAE, one patient for anti- Mi-2 and four patients for anti-TIFγ. Additionally, two patients were negative for MSAs. The clinical features of the patients, including MSA status, skin lesion, ILD, oesophageal lesions and muscle disease, are displayed in table 1.
Clinical characteristics of patients with DM
β5i and RIG-I expression levels increased significantly in DM-affected muscle tissue
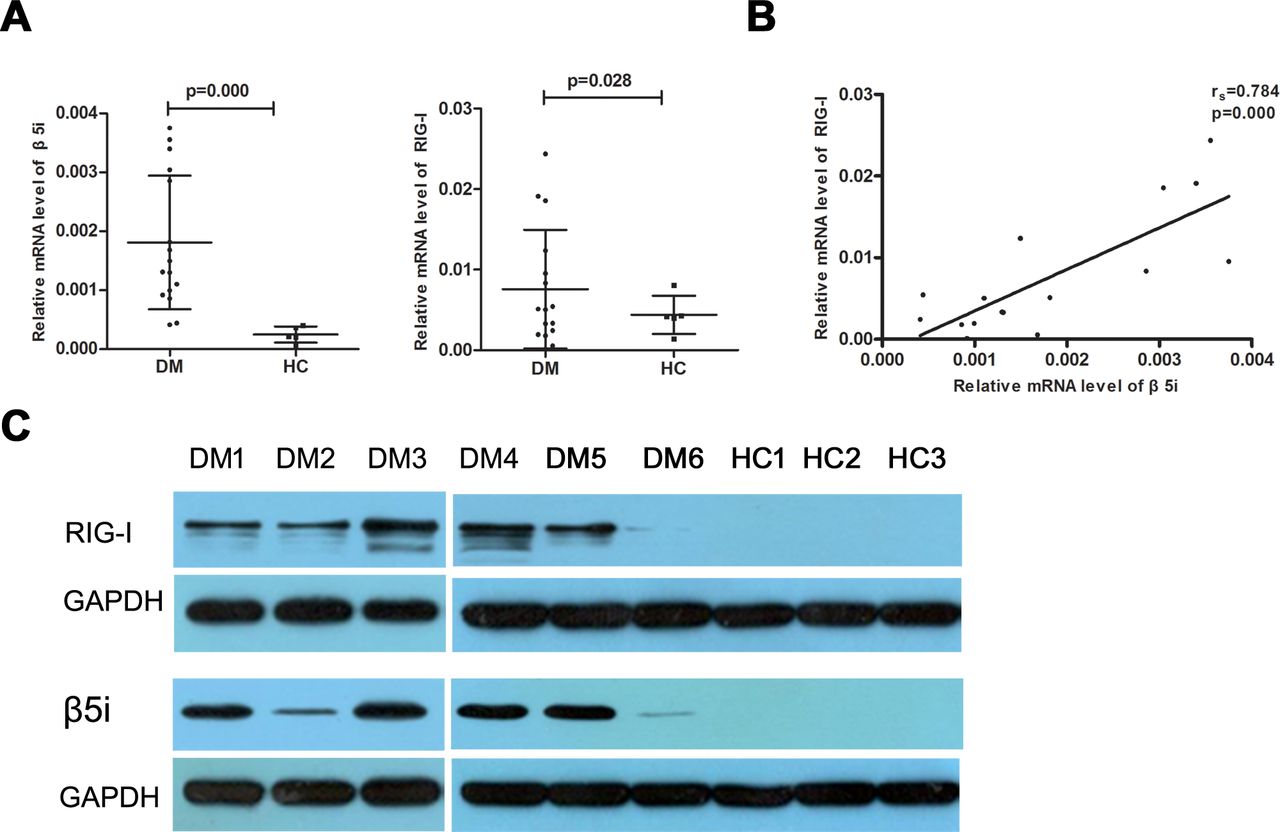
The real-time RT-PCR analysis revealed significantly higher β5i and RIG-I mRNA expression levels (0.0018±0.0011 vs 0.0003±0.0001, p=0.000 and 0.0076±0.0074 vs 0.0029±0.0012, p=0.028, respectively) in the muscle tissue of patients with DM than those in the controls (figure 1A). Meanwhile, significant positive correlations were observed between β5i and RIG-I mRNA levels (rs=0.784, p=0.000; figure 1B). In addition, patients with different MSAs had various level of β5i and RIG-I mRNA; however, we failed to analyse the statistical significance because of the small sample size in each MSA subgroup (online supplemental figure 1). The β5i and RIG-I proteins were detected in the muscle tissues of six patients with DM and three controls by western blotting. As shown in figure 1C, β5i and RIG-I expression in patients with DM was significantly higher than that in the controls. Further, we analysed the expression of β5i and RIG-I by immunohistochemistry; β5i and RIG-I were more prominently expressed in the perifascicular areas of muscle fibres while no obvious staining was found in controls (figure 2A–F). In addition, the double-immunofluorescence staining analysis showed the notable colocalisation of β5i and RIG-I in the perifascicular regions (figure 2G).
Supplemental material
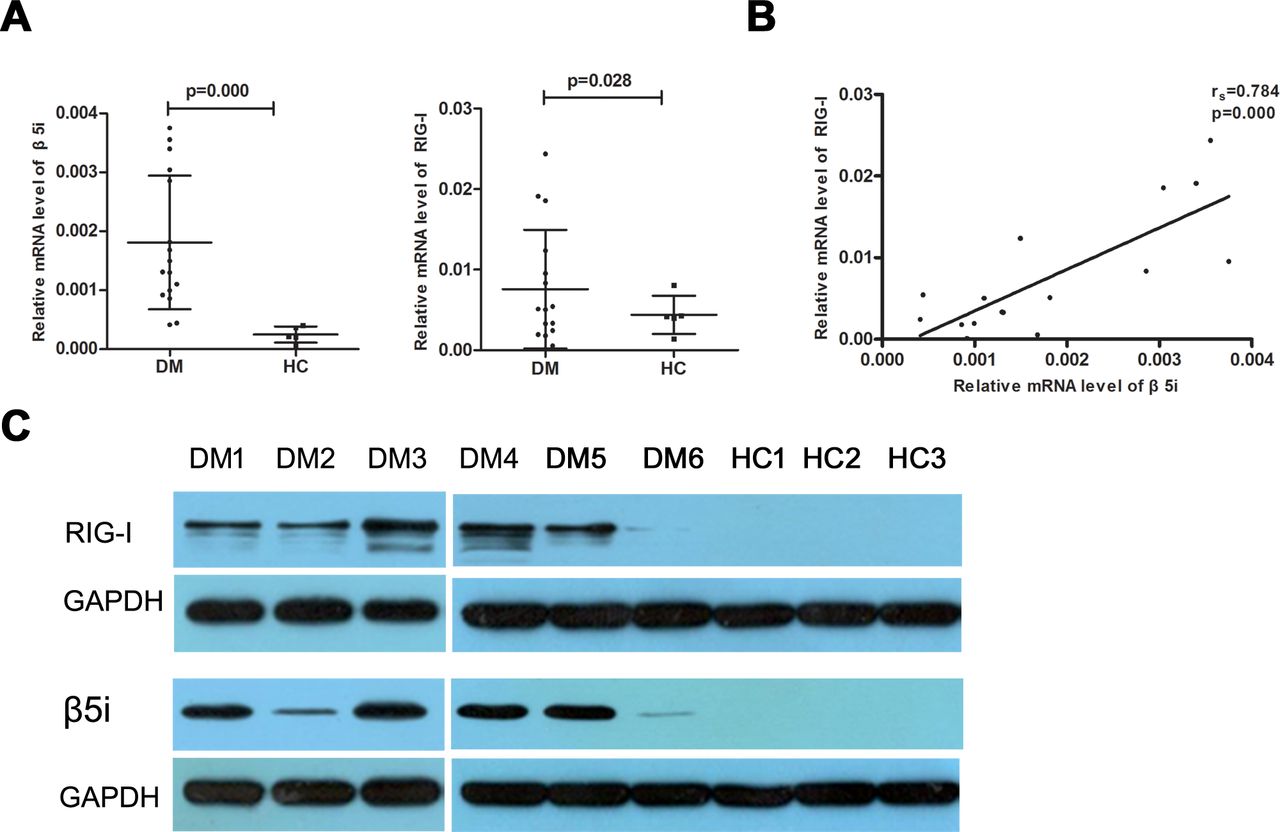
β5i and RIG-I expression levels increased significantly in DM affected muscle tissue. (A) Patients with DM exhibited significantly higher levels of β5i and RIG-I mRNA than controls (HC). (B) The β5i mRNA level correlated positively with RIG-I mRNA level. (C) Western blotting analysis of β5i and RIG-I in six DM and three controls muscle. DM, dermatomyositis. HC. healthy control.
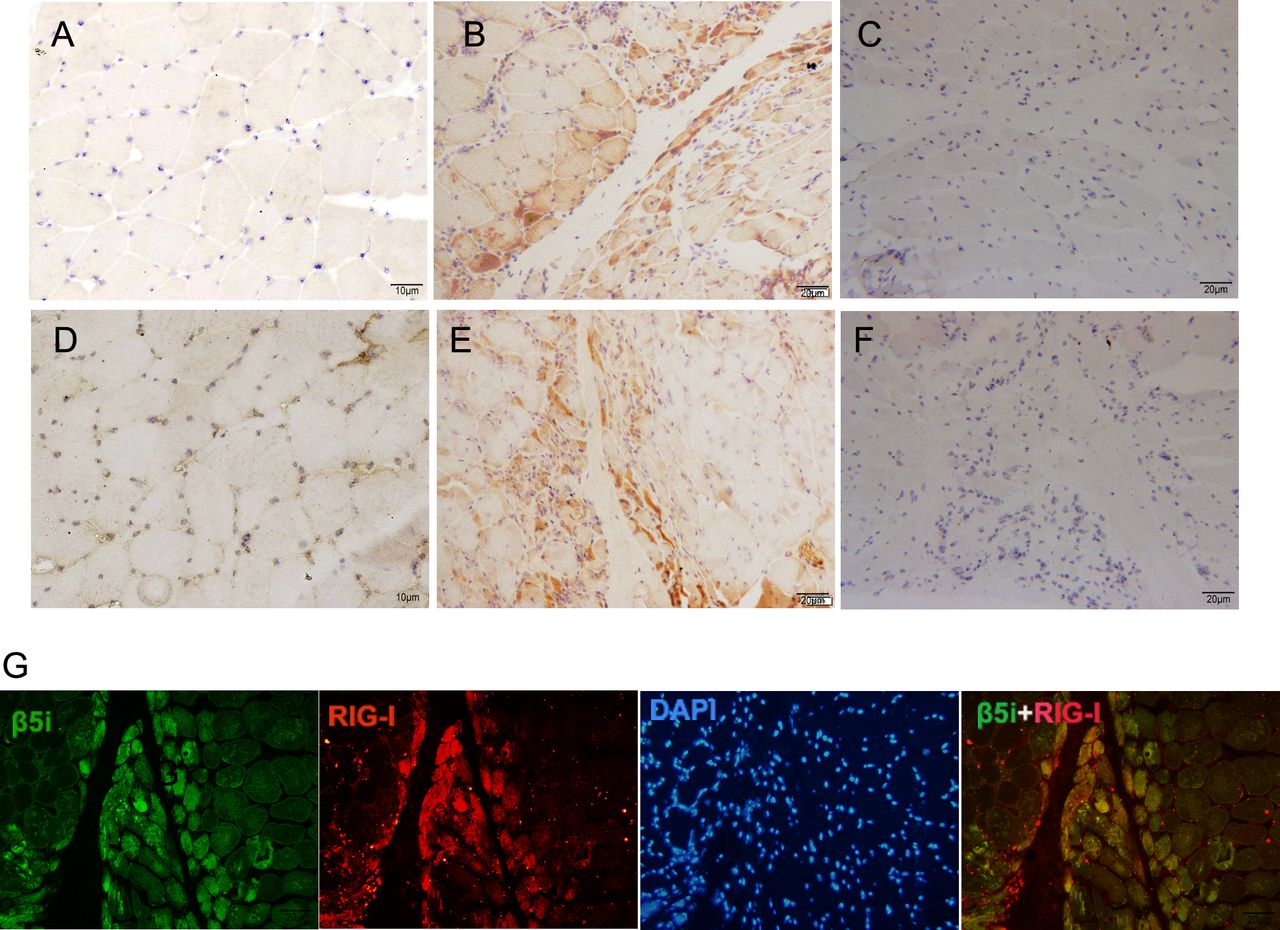
High expression of β5i and RIG-I colocalised in the perifascicular myofibres of DM. (A) Immunohistochemistry staining of β5i in muscle from a control subject. (B) Immunohistochemistry staining of β5i in muscle from a patient with DM. (C) A recombinant monoclonal rabbit antibody-isotype control for β5i staining of muscle from a patient with DM. (D) Immunohistochemistry staining of RIG-I in muscle from a control subject. (E) Immunohistochemistry staining of RIG-I in muscle from a patient with DM. (F) A polyclonal rabbit antibody-isotype control for RIG-I staining of muscle from a patient. (G) Double immunofluorescence staining show β5i (green) and RIG-I (red) colocalised in the perifascicular myofibres of DM. DM, dermatomyositis.
β5i and RIG-I expression was closely related to muscle disease severity
The β5i and RIG-I mRNA expression levels in muscle tissues were associated with muscle VAS (rs=0.601, p=0.014 and rs=0.524, p=0.037, respectively) and MMT8 score (rs =−0.561, p=0.024 and rs =−0.464, p=0.070, respectively), but did not correlate with serum CK levels (table 2). As for the muscle pathological score, the β5i and RIG-I mRNA levels were positively correlated with the muscle fibre domain score (rs=0.533, p=0.033, and rs=0.731, p=0.001, respectively; table 2).
The correlation between ß5i./RIG-I mRNA level and skeletal muscle disease
β5i is not expressed in regenerative myocytes
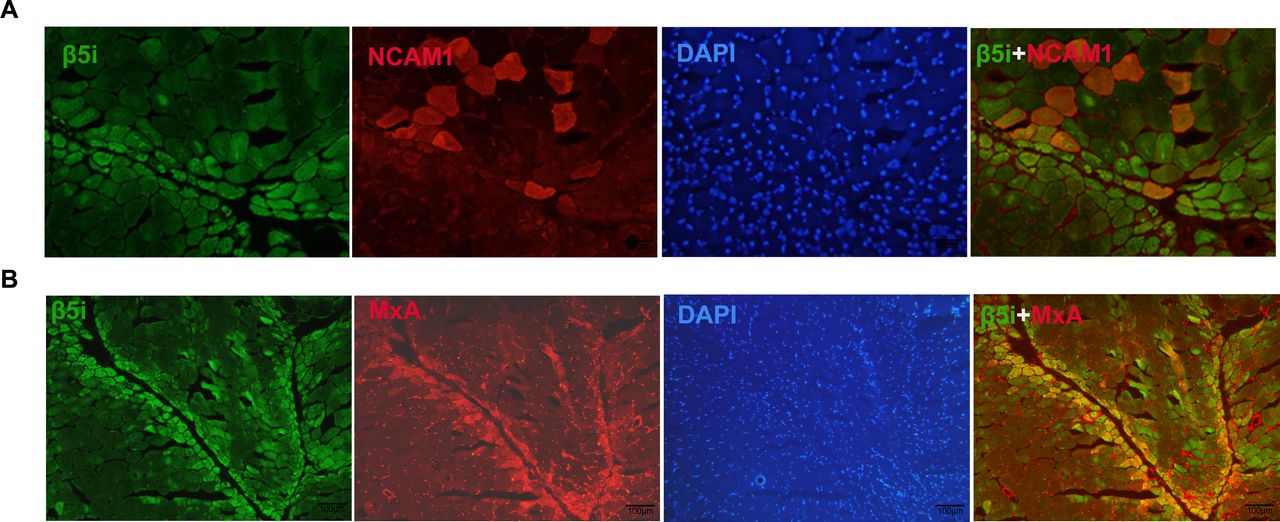
Immunohistochemistry analyses revealed that β5i was located mainly in the perifascicular regions of small myocytes (figures 2B and 2E). We performed a double-immunostaining assay to test for the presence of NCAM1, a regenerative cell marker. As shown in figure 3A, β5i was mainly located in NCAM1-negative myocytes. Furthermore, we performed double-label immunofluorescence staining of β5i and MxA. The results showed the colocalisation of β5i and MxA in the perifascicular atrophy region (figure 3B).

β5i is not expressed in regenerative myocytes. (A) Immunofluorescence staining show β5i (green) mainly located in NCAM1 (red) negative myocytes. (B) Double immunofluorescence staining of β5i and MxA show that the colocalisation of β5i and MxA in the perifascicular atrophy region.
Overexpression of β5i activates the NF-κB pathway, promotes RIG-I expression and causes myoblast atrophy
To investigate the potential links between β5i and muscle atrophy, we overexpressed β5i in HSMMs and consequently noted the upregulation of MxA, IFN-β and RIG-I (figure 4A,B). Meanwhile, upregulation of phosphorylated IKBα (pIκBα), IRF3 (pIRF3) and NF-κB (pNF-κBp65) in β5i-overexpressing HSMMs suggested that β5i may activate the NF-κB pathway (figure 4A,B). Interestingly, we found that the expression of the MuRF1 protein was also increased in β5i-overexpressing HSMMs (figure 4A,B). In addition, β5i inhibition with PR957 significantly attenuated the upregulation of MxA, IFN-β, RIG-I, MURF1, pIKBα, pIRF3 and pNF-κBp65 (figure 4). The viability of β5i-overexpressing HSMMs decreased significantly compared with that of the HSMMs from the control group (1.09±0.16 vs 2.64±0.27; p=0.000); this decrease was partially recovered by treatment with PR957 (figure 4C,D).

Overexpression of β5i upregulate NF-κB pathway related markers, promote the RIG-I expression and myoblast atrophy. (A) Protein expression of β5i, MxA, IFN-β, RIG-I, MURF1, pIκBα, pIRF3 and pNF-κB in different HSMMs (Control, β5i overexpression and β5i OE with PR957) by Western Blot analysis. (B) Bar graph shows the quantification of protein expression relative to β-actin level. Data expressed as mean±SD (n=3 per group). (C) HSMMs were observed for morphological changes at 24 hours after treatment (×200). (D) The HSMMs viability was inhibited significantly after β5i OE and partly recovered by treatment with PR957. CCK-8 method was used to examine cell viability and the optical density (OD) values at 450 nm were measured by microplate reader. *p<0.05 vs control. #p<0.05 vs β5i OE. HSMMs, human skeletal muscle myoblasts; OE, overexpression.
Activation of RIG-I by pppRNA elevated IFN expression and promoted myoblast atrophy
To investigate the effects of RIG-I activation on myoblast atrophy, an RIG-I specific ligand pppRNA was used to stimulate HSMMs. In response to pppRNA stimulation, the expression levels of RIG-I, MxA, IFN-β and MURF1 increased (figure 5A). In addition, activation of RIG-I by pppRNA caused a marked reduction in HSMM viability within 24 hours, compared with the viability of HSMMs in the control group (0.81±0.09 vs 0.62±0.05; p=0.001; figure 5B,C).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Activation of RIG-I by pppRNA elevated IFN and MURF1 and reduced viability of HSMMs. (A) Protein expression of RIG-I, MxA, IFN-β and MURF1 were upregulated in myoblast after pppRNA 24 hours activation. The intensity values of Western blot were measured with Image J software and intensity values were measured compared with the respective β-actin which are shown below the band. (B) HSMMs were observed for morphological changes at 24 hours after treatment (×200). (C) The HSMMs viability was inhibited significantly after pppRNA 24 hours activation. CCK-8 method was used to examine cell viability and the optical density (OD) values at 450 nm were measured by microplate reader. HSMMs, human skeletal muscle myoblasts.
Discussion
In this study, we identified the high, colocalised expression of β5i and RIG-I in perifascicular myofibres in DM. Moreover, the expression of β5i and RIG-I in muscle tissue was closely related to muscle disease severity. Furthermore, in vitro experiments revealed that β5i overexpression in HSMMs activates the NF-κB pathway and promotes RIG-I upregulation, decreasing the viability of HSMMs. Thus, β5i and RIG-I may contribute to the pathogenesis of the DM-associated perifascicular atrophy of muscles.
Previous studies have demonstrated the upregulation of β5i and RIG-I in the muscles in patients with DM.8 10 20 21 Further, Suárez-Calvet et al reported that positive RIG-I staining in perifascicular myofibres is a reliable biomarker for the diagnosis of DM.22 Interestingly, we found that the upregulation of β5i and RIG-I expression in muscle was closely related to the clinical severity of muscle injury and the pathological score of muscle fibres. Therefore, our data suggest that β5i and RIG-I may be potential biomarkers of muscle damage in DM.
The pathogenesis of DM is poorly understood. Perifascicular myofibre atrophy is a major pathological hallmark of DM. Although it is well known that marked overproduction of type I IFN inducible transcripts and proteins predominate in the perifascicular myofibres of DM-affected muscles,2 3 research regarding the mechanisms underlying DM-associated muscle impairment is limited. However, an in vitro study showed that type I IFN might induce myoblast atrophy and vascular disruption in DM.23 Another study has suggested that IFN-β induced reactive oxygen species production in human myotubes, causing mitochondrial malfunctions and ultimately leading to muscle damage.24 Our data showed that RIG-I activation by pppRNA upregulated IFN expression and triggered myoblast atrophy, providing new evidence that RIG-I is a key regulator involved in the mechanisms underlying DM-associated muscle damage.
Proteasomes are large multisubunit protein complexes responsible for most of the non-lysosomal proteolysis occurring in cells.25 The standard 20S proteasome comprises three subunits, β1, β2 and β5. On stimulation with cytokines, such as IFN-γ, the formation of three alternative subunits, β1i, β2i and β5i, is induced; these replace the standard subunits to form the 20S immunoproteasome.26 The immunoproteasome exerts various biological functions, including the improvement of MHC-I antigen presentation, proinflammatory cytokine production, T cell differentiation and survival and defence against oxidative stress.27–29 Previous studies have shown that the expression of β5i significantly increased both in perifascicular myofibres and muscle infiltrating cells (CD8+ T-cells and CD68+ macrophages), which are involved in influencing the pathological MHC-I expression, as well as the regulation of proinflammatory cytokine responses in muscle tissues.10 20 In addition, β5i may promote the activation of NF-κB in podocytes of animal models of diabetes and in murine tissues of aortic aneurysms.30 Animal models of deficient β5i expression have been reportedly associated with the attenuation of colitis due to the reduced expression of many NF-κB-regulated proinflammatory mediators.31 In this study, we noted an elevation in β5i expression, which was predominant in the perifascicular area and closely related to skeletal muscle disease severity. To assess the relationship between β5i expression and myoblast atrophy, we overexpressed β5i in vitro. We found that β5i overexpression upregulated the NF-κB pathway-related markers and promoted type I IFN and RIG-I expression, as well as the muscle atrophy related marker MuRF1. Meanwhile, we observed that the β5i-overexpressing HSMMs grew slowly and showed decreased viability, as confirmed by the CCK8 test. Decreased cell viability combined with increased MuRF1 expression indicated HSMMs atrophy. Thus, our data suggest that β5i may contribute to myoblast atrophy by promoting the upregulation of type I IFN and RIG-I.
In addition, as an important member of the RLRs family, RIG-I can recognise and bind to double-stranded viral RNA molecules; in turn, it enhances the transcription of type I IFN.32 33 On the other hand, type I IFN may also upregulate RIG-I.33 34 The marked elevation in the expression levels of RIG-I and type I IFN in perifascicular atrophy area suggests that abnormal positive feedback between RIG-I and type I IFN influences the pathogenesis of DM. Our data showed that β5i upregulated type I IFN and RIG-I by activating the NF-κB pathway. Therefore, β5i overexpression may be a possible factor underlying sustained type I IFN overproduction in DM.
We acknowledge that this study has a few limitations. First, the immunoproteasome comprises three subunits: β1i, β2i and β5i; however, we did not detect the expression of β1i and β2i in our research. A study by Bhattarai et al showed that the staining of β1i was also more prominent in the perifascicular area in DM-affected muscles.10 Whether β1i and β2i also participate in perifascicular atrophy warrants further investigation. Furthermore, chymotrypsin-like (CT-L) activity is an important proteolytic function of β5i. We only detected the elevated expression of β5i but did not assess its CT-L activity. On this basis, future evaluation of the CT-L activity may help clarify the mechanism underlying the effects of β5i on myoblast atrophy. Third, MSAs have been extensively demonstrated to correlate with specific clinical phenotypes. We found patients with different MSAs have various levels of β5i and RIG-I mRNA. However, the number of subgroup cases was too small to analyse the statistical significance, which should be further studied based on larger sample size. Finally, the role of RIG-I in myoblast atrophy was only studied by pppRNA-mediated RIG-I stimulation, and whether β5i can directly upregulate MuRF1 and prompt myoblast atrophy needs to be further studied. We believe that further in-depth studies with RIG-I-knockout myoblasts can be more reliable in verifying the role of RIG-I in β5i-mediated myoblast atrophy.
In conclusion, our data suggest a novel role of the immunoproteasome subunit β5i as a critical regulator that promotes perifascicular atrophy in DM. Our findings provide insights into the mechanisms underlying DM-associated muscle damage and underscore a new potential therapeutic target for DM.
Data availability statement
Data are available on reasonable request.
Ethics statements
Patient consent for publication
Ethics approval
This study involves human participants and was approved by Ethics Committee of China Japan Friendship Hospital (2019-25-K19). Participants gave informed consent to participate in the study before taking part.
Acknowledgments
We would like to thank Editage (www.editage.cn) for English language editing.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Footnotes
LZ and QX contributed equally.
Contributors All the authors contributed to the manuscript, conception and design, collection of data, analysis and interpretation. All the authors reviewed the final version of the manuscript. LZ, QX and QP contributed to study conception, study design, data acquisition, data analysis, data interpretation and drafting and revising the manuscript. QX and LY conducted cell experiments, molecular biology experiment, PCR analysis and Western blotting analysis. WL and QL conducted Immunohistochemistry and Immunofluorescence. LZ and XT recruited patients and participated in data recording and clinical and muscle pathology evaluation assessment. GW helped in data interpretation and revised the manuscript. QP is responsible for the overall content as guarantor. All authors read and approved the final manuscript.
Funding This work was supported by the National High Level Hospital Clinical Research Funding (2022-NHLHCRF-YS-02),National Natural Science Foundation of China (81601425, 82171788) and the Elite Medical Professionals project of China-Japan Friendship Hospital (NO. ZRJY2021-GG13).
Competing interests None declared.
Provenance and peer review Not commissioned; externally peer reviewed.
Supplemental material This content has been supplied by the author(s). It has not been vetted by BMJ Publishing Group Limited (BMJ) and may not have been peer-reviewed. Any opinions or recommendations discussed are solely those of the author(s) and are not endorsed by BMJ. BMJ disclaims all liability and responsibility arising from any reliance placed on the content. Where the content includes any translated material, BMJ does not warrant the accuracy and reliability of the translations (including but not limited to local regulations, clinical guidelines, terminology, drug names and drug dosages), and is not responsible for any error and/or omissions arising from translation and adaptation or otherwise.