Article Text

Abstract
Objective Mast cells may play a role in rheumatoid arthritis (RA), but activation of human mast cells in autoimmune settings has been little studied. Toll-like receptors (TLR) and Fcγ receptors (FcγR) are important receptors for cellular activation in the joint, but expression and stimulation of these receptors in human mast cells or the functional interplay between these pathways is poorly understood. Here, we analysed triggering of human mast cells via these receptors in the context of anti-citrullinated protein antibody-positive (ACPA+) RA.
Methods RNA and protein expression of TLRs and FcγR was quantified using PCR and flow cytometry, respectively. Mast cells were stimulated with TLR ligands (including HSP70) combined with IgG immune complexes and IgG-ACPA.
Results Human mast cells expressed TLRs and produced cytokines in response to TLR ligands. Both cultured and synovial mast cells expressed FcγRIIA, and triggering of this receptor by IgG immune complexes synergised with activation by TLR ligands, leading to two- to fivefold increased cytokine levels. Mast cells produced cytokines in response to ACPA immune complexes in a citrulline-specific manner, which synergised in the presence of HSP70.
Conclusions Our data show that synovial mast cells express FcγRIIA and that mast cells can be activated by IgG-ACPA and TLR ligands. Importantly, combined stimulation via TLRs and immune complexes leads to synergy in cytokine production. These findings suggest mast cells are important targets for TLR ligands and immune complexes, and that combined activation of mast cells via these pathways greatly enhances inflammation in synovial tissue of RA patients.
- Autoantibodies
- Rheumatoid Arthritis
- Ant-CCP
Statistics from Altmetric.com
Introduction
Rheumatoid arthritis (RA) is an autoimmune disease characterised by chronic inflammation of the synovial lining of the joint. Antibodies against citrullinated proteins (ACPA) can be found in the majority of patients with established RA.1 These antibodies recognise proteins in which arginine is converted into citrulline through a post-translational modification. ACPA are associated with disease progression2 and their pathogenic potential has been shown in mice.3 ,4 The exact way by which ACPA contribute to inflammation is not known, but ACPA immune complexes have been shown to allow the activation of monocytes and macrophages.5 ,6 However, several other immune cells may contribute to ACPA-mediated immune responses in RA.
One of these cell subsets are mast cells, which comprise up to 5% of the synovial cells. Several findings in patients support an active role of mast cells in RA pathogenesis. Mast cell numbers are increased in RA synovium.7 ,8 Furthermore, several mast cell mediators have been observed in the synovial fluid of RA patients,8 ,9 and these mediators have been shown to have pathogenic effects on synovial fibroblasts,10 ,11 suggesting that mast cells may play a role in inflammatory processes in RA. However, the specific pathways leading to mast cell activation in RA have not been shown.
The main biological function of mast cells is considered to be protection against pathogens.12 Recognition of pathogens by mast cells via toll-like receptors (TLRs) may lead to inflammatory responses such as recruitment of neutrophils and T cells. TLR ligands can also be released during chronic inflammatory responses, and are thought to contribute to the chronicity of RA.13–17
Although studies performed in mice suggest that mast cells can be activated by TLR ligands, there is only limited information on TLR triggering of human mast cells. Some studies have shown expression of TLR-2, -3 and -4 by human mast cells, but the response to other TLRs and the type of response that is induced in mast cells is not clear.18–22
It has been suggested that the innate responses of mast cells towards pathogen-derived or endogenous TLR ligands may be enhanced by the presence of specific antibodies, such as those present during a memory response to pathogens, allowing a rapid clearance of the infecting microbe.12 However, in the context of autoimmunity, such an enhanced response can have harmful effects, and may contribute to aberrant and continuous activation of the immune system.
As mast cells are present in RA synovium, a location where endogenous TLR ligands and citrullinated proteins have been found, combined mast cell activation via TLR and ACPA immune complexes could play a role in chronic immune activation in RA. Because the expression of TLRs and the functional response towards TLR ligands in human mast cells is not established, and because it is not known whether mast cells can be activated by ACPA immune complexes, we aimed to thoroughly analyse mast cell responses upon TLR and FcγR triggering as well as the interplay between TLR- and IgG-mediated responses in ACPA+ RA.
Methods
Peripheral blood-derived mast cells
Buffy coats from healthy volunteers were obtained from a blood bank (Sanquin, The Netherlands). CD34+ haematopoietic stem cells were isolated from peripheral blood mononuclear cells (PBMCs) with CD34 microbeads (Miltenyi Biotec). Isolated stem cells were differentiated into mast cells as described.23 After 6–8 weeks, the purity of mast cells was determined by flow cytometric analysis of CD117, FcεRI, CD203c and intracellular tryptase. The purity of mast cells ranged from 90% to 99%.
Real-time PCR
One million mast cells per donor were lysed in TRIzol reagent (Invitrogen). RNA was isolated using TRIzol. A cDNA library was constructed using the RT2 First Strand Kit (SA Biosciences). Expression of mRNA for TLRs and the TLR signalling pathway was assessed by real-time PCR, using the Toll–Like Receptor Signaling Pathway PCR Array (SA Biosciences). Expression was normalised to housekeeping gene HPRT1.
Mast cell activation
Cultured mast cells were sensitised with 0.1 μg/mL IgE (Diatec) for 18 h, after which they were activated with 10 μg/mL goat anti-human IgE (Nordic). For TLR-mediated activation, mast cells were stimulated using 1 μg/mL Pam3csk, 10 μg/mL zymosan, 10 μg/mL Staphylococcus aureus peptidoglycan, 10 μg/mL poly(I:C), 0.1–10 μg/mL Escherichia coli K12 lipopolysaccharide (LPS), 1 μg/mL Salmonella typhimurium flagellin, 1 μg/mL FSL-1, 5–10 μg/mL imiquimod, 10 μg/mL ssRNA40 and 5 μM ODN2006 (CpG) (all from InvivoGen) for 24 h. For FcγR-mediated activation by plate-bound IgG (pb IgG), culture plates (U-bottom, 96 wells) were coated with 10 μg/mL purified human IgG (Jackson ImmunoResearch) in PBS at 37° for 1.5 h.24 For activation by soluble immune complexes, human IgG was heat aggregated (HA) at 63°C for 30 min, after which insoluble complexes were removed by centrifugation at 13 000 rpm for 5 min. Mast cells were stimulated using 100 μg/mL HA IgG.25
For blocking studies, mast cells were incubated with 20 μg/mL anti-CD32 (clone IV.3; Stemcell), mIgG2b isotype control (eBioscience), 10 μg/mL anti-TLR-2 (clone TL2.1; eBioscience) or anti-TLR-4 (clone HTA-125; eBioscience) for 1 h at 37° prior to activation, after which cells were diluted 1:1 in culture medium containing the indicated stimuli.
IgG-ACPA mediated mast cell activation
Biotinylated CCP2 peptide and the arginine control peptide were obtained from Dr J W Drijfhout, Department of Immunohematology and Bloodtransfusion, LUMC, Leiden, The Netherlands. Nunc MaxiSorp plates (VWR) were first coated overnight with 5 μg/mL streptavidin (Sigma). After washing, biotinylated CCP2 and arginine control peptides (1 μg/mL) were coated for 1 h at room temperature, and the coated wells were subsequently incubated for 1 h at 37°C with serum of ACPA-positive and ACPA-negative RA patients, diluted 50× in PBS/0.1% BSA. After washing, mast cells were added, and were cultured in the presence or absence of 10 μg/mL human HSP70 (Enzo Life Sciences). Alternatively, CCP2- or arginine control-coated plates were incubated with 5 μg/mL ACPA mAb. The ACPA IgG monoclonal antibody used here was previously cloned by van de Stadt et al.26 Briefly, ACPA antibody was produced following transduction with heavy and light chain cDNA expressing lentiviral vectors by HEK 293 T cells,27 and antibodies were purified from supernatant using fast protein liquid chromatography (ÄKTA; GE Healthcare) equipped with HiTrap Protein A columns (GE Healthcare).28 Following purification, the quantity of ACPA was assessed by standard total IgG ELISA (Bethyl Laboratories) according to the manufacturer's instructions.
Patient samples
Diagnoses of RA or osteoarthritis (OA) were made according to 1987 and 1986 American College of Rheumatology criteria, respectively.29 ,30 The presence of total IgG-ACPA and rheumatoid factor (RF) in serum was tested by routine diagnostic ELISA.
Synovial tissues were obtained from patients with established RA and OA who had undergone knee or hip replacement surgery. Synovial tissue was digested in 1 mg/mL collagenase IA (Sigma-Aldrich) and 50 μg/mL DNAse (Sigma–Aldrich) at 37°C for 1 h, after which single cell suspensions were obtained using a 70 μm filter. All patients gave written informed consent, and the study was approved by the Leiden University Medical Center human ethics committee.
Flow cytometry
A list of antibodies used in this study is shown in online supplementary table S1.
For surface staining, cells were incubated with fluorochrome-conjugated antibodies diluted in PBS 0.5% BSA at 4°C for 30 min. Intracellular tryptase staining was performed as described.23 Flow cytometric acquisition was performed on FACSCalibur (BD) and LSR II (BD) flow cytometers. Analysis was performed using FACSDiva (BD) and FlowJo software.
Cytokine and leukotriene C4 production
Quantitative immunoassays in culture supernatants were performed using the 42-plex cytokine Milliplex assay (Millipore). Additionally, IL-8 production was evaluated using ELISA (eBioscience). Leukotriene C4 was analysed using a competitive ELISA kit (Neogen).
Statistical analysis
Results are expressed as mean±SEM. For detection of TLR mRNA expression, the one-sample t test was performed. For differences between multiple groups, one-way ANOVA was performed, with Bonferroni's post hoc test to correct for multiple testing. Synergy was defined as an increase in mast cell activation with the combination of stimuli compared to the additive mast cell response induced by the stimuli separately, determined using the paired-samples t test. p Values of <0.05 were considered statistically significant.
Results
Expression of TLR
To determine the expression of TLR1 to -10 and other molecules involved in TLR signalling pathways, real-time PCR was performed on the mRNA of peripheral blood-derived mast cells. Mast cell cultures were defined by expression of CD117, CD203c, FcεRI and intracellular tryptase, indicating that the culture resulted in mature mast cells (see online supplementary figure S1). mRNA encoding TLR-1 to -9 was found in all three mast cell donors (figure 1A), although expression of TLR-3 and -9 was low compared to the other TLRs. The relative expression levels of TLRs in mast cells were comparable to PBMCs and most often higher than in the monocyte cell line THP-1 (see online supplementary figure S2). Mast cells also expressed mRNA for several molecules associated with TLR signalling, such as MyD88, TICAM1 (TRIF), TICAM2 and BTK (figure 1B).

Total mRNA was isolated from independent peripheral blood-derived mast cell cultures of different donors (n=3). Relative mRNA expression of (A) Toll-like receptors (TLRs) and (B) molecules involved in TLR signalling, normalised to HPRT1 expression as housekeeping gene. The dotted line indicates the detection limit. nd, not detected.
These data indicate that human mast cells express TLR-1 to -9 as well as the downstream signalling molecules, needed for TLR signal transduction.
Mast cell activation by TLR ligands
To examine whether the expression of TLRs by mast cells could be related to functional responses, cytokine production in response to TLR ligands was measured. Figure 2A and online supplementary figure S3 show that mast cells respond to TLR ligands with the production of several cytokines. Cytokine responses to ligands for TLR-3 and -9 were low or absent, in line with their low mRNA expression. In contrast, GM-CSF, IL-8 and MIP-1α were produced in response to most TLR ligands, IL-10 and IL-13 were mainly produced in response to ligands for TLR-2, and TNF-α production was mainly restricted to ligands for TLR-2, -4 and -8.

(A) Mast cells were stimulated with the indicated Toll-like receptor (TLR) ligands for 24 h after which cytokines were measured in supernatant. The stimulation index was calculated by dividing the amount of cytokine produced after stimulation by the amount of cytokine in the supernatant of unstimulated mast cells. (B–D) Mast cells were stimulated with TLR ligands for 1 h, after which cells were used for flow cytometry to assess degranulation with anti-CD63 (B,C) and supernatant was used for the determination of leukotriene C4 (LTC4) release (D). Data were obtained from two independent experiments performed in duplicate. Asterisks indicate significant increases compared to unstimulated mast cells (p<0.001).
Besides cytokine production, one of the main effector functions of mast cells is mediated via granule release and production of lipid-derived mediators. In contrast to stimulation with anti-IgE, these responses were absent when mast cells were activated with TLR ligands (figure 2B–D). Therefore, TLR ligands induce a different response as compared to anti-IgE, characterised by the production of several cytokines in the absence of degranulation or release of lipid-derived molecules. Importantly, different TLR triggers can induce different response modes as exemplified by different cytokines induced by different TLR ligands.
Activation of mast cells by IgG immune complexes via FcγRII
As the expression of FcγRs by human mast cells and their possible activation via these receptors is not established, we next evaluated the potential of human mast cells to be activated by IgG immune complexes. First, we evaluated expression of the different FcγRs by flow cytometry (figure 3A,B). Mast cells were found to express FcγRIIA (CD32A) but no other FcγR. We next stimulated mast cells with IgG immune complexes, and found that mast cells produced IL-8 in response to plate-bound IgG (figure 3C). Importantly, this response was almost completely inhibited by blocking antibodies to FcγRIIA (figure 3D). In addition, mast cells produced IL-8 in response to soluble immune complexes (HA IgG), and this response was also blocked by anti-FcγRIIA (figure 3E). No degranulation was observed in response to IgG immune complexes (data not shown). Together, these data show that human mast cells are activated by IgG immune complexes, mainly via FcγRIIA.
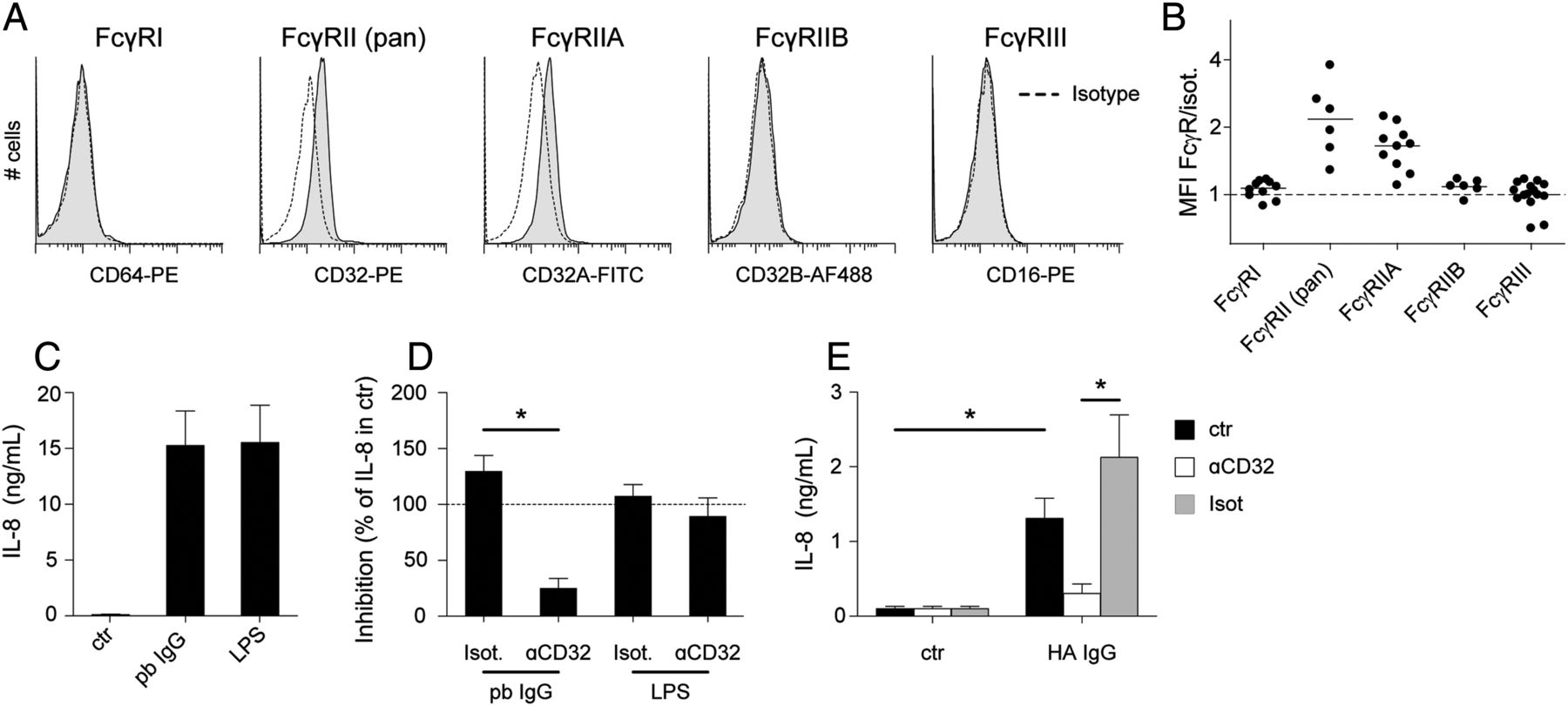
(A,B) Expression of Fcγ receptors (FcγR) by cultured mast cells was analysed by flow cytometry. Representative examples are shown in (A) and a summary of all donors (n=6–15) analysed is shown in (B). (C) IL-8 production by mast cells in response to plate-bound (pb) IgG and lipopolysaccharide (LPS) (n=13). (D) Inhibition of IL-8 production in response to pb IgG or LPS by anti-CD32 or isotype control (n=6), indicated as a percentage of total IL-8 production in stimulated mast cells without blocking antibodies. (E) IL-8 production in response to heat-aggregated IgG (HA IgG), and inhibition by anti-CD32 or isotype control (n=4).
Synergy between TLR- and FcγR-mediated activation
As mast cells responded to triggering of TLR as well as FcγRII, we next evaluated the possible interaction between triggering with IgG immune complexes and TLR ligands (figure 4A,B). No additional effects were observed when TLR-3 or -9 ligands were combined with plate-bound IgG, which is in line with the absence of activation with these ligands. However, combination of other TLR ligands with IgG led to synergy in cytokine production (figure 4B). Similar to the results obtained with plate-bound IgG, IL-8 production by mast cells in response to LPS was greatly enhanced by soluble immune complexes (figure 4C).


Mast cells were stimulated with Toll-like receptor (TLR) ligands alone or in combination with plate-bound IgG immune complexes. (A–C) IL-8 production was measured in supernatant after 24 h upon combined triggering with plate-bound (pb) IgG and TLR ligands. (A) Summary graphs showing the mean±SEM of IL-8 in supernatant (n=3). (B) IL-8 production for all TLR ligands in combination with plate-bound IgG is shown (closed symbols) compared to the sum of cytokine levels following stimulation with IgG and TLR ligands separately (open symbols). Each symbol represents an independent donor. Asterisks indicate significantly (p<0.05) increased cytokine production when TLR ligands were combined with anti-IgE compared to the cumulative effect of the stimuli alone. (C) IL-8 production in response to heat-aggregated IgG (HA IgG) in combination with lipopolysaccharide (LPS) (n=4). (D) IL-8 production in response to plate-bound IgG in combination with zymosan (zym) and LPS after blocking with anti-CD32 or isotype control (n=2).
Expression of FcγRs was not changed by TLR ligands (data not shown), indicating that the synergistic effect was dependent on FcγRIIA, the only FcγR that was expressed. Indeed, blocking of FcγRIIA led to almost complete inhibition of IL-8 production, resulting in IL-8 production comparable to the TLR ligands alone (figure 4D). These results indicate that combined stimulation of mast cells via TLR and FcγRIIA can markedly enhance inflammatory responses by mast cells.
Expression of FcγR by synovial mast cells
Although mast cells are present in synovial tissue, the number of cells that can be isolated in a non-activated manner does not allow for in vitro mast cell activation for functional studies. Nonetheless, we were able to analyse the expression of FcγR by synovial mast cells by flow cytometry (figure 5). Synovial mast cells from all RA and OA donors were found to express FcγRII. Further analysis in three donors showed that FcγRIIA, and not IIB, is expressed by synovial mast cells. Expression of other FcγRs was very low or absent, except for one donor, in which both FcγRI and FcγRIII were detected. These results confirm the comparability between cultured mast cells and synovial mast cells regarding FcγR expression and indicate that the molecular make-up of synovial mast cells allows them to be activated by IgG immune complexes via FcγRIIA.

(A) Representative example of mast cell gating is shown. Mast cells were gated as CD117+IgE+ or CD117+FcεRI+ cells within the live cells as gated based on forward (FSC) and sideward scatter (SSC). Representative examples of Fc receptor staining are shown in (B) and a summary of all patients analysed (n=6 rheumatoid arthritis patients and n=4 osteoarthritis patients) is shown in (C), with each symbol representing an individual patient.
Synergy between TLR- and IgG-ACPA-mediated activation of mast cells
We next wished to evaluate whether human mast cells can respond to ACPA immune complexes to confirm the observed synergy between IgG- and TLR-mediated activation in an antigen-specific system.
First, mast cells were stimulated with ACPA immune complexes on CCP2-coated plates, resulting in IL-8 production (figure 6A,B). This activation was citrulline dependent, as the arginine control peptide, or the use of serum from ACPA-negative patients, did not induce IL-8 release. Mast cell activation was inhibited by pre-incubating mast cells with an anti-FcγRIIA antibody (figure 6C). The use of RF-negative serum (figure 6D) or an ACPA monoclonal IgG antibody (figure 6E) also induced IL-8 production, confirming that mast cell activation by ACPA immune complexes is not dependent on the presence of RF. These results show that mast cells can be activated by ACPA immune complexes, and that this is mainly mediated via FcγRIIA.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
(A, B) Mast cells were stimulated on CCP-coated plates incubated with no serum (ctr) or serum of antibodies against citrullinated proteins (ACPA+) and ACPA− rheumatoid arthritis patients, as indicated, for 24 h, after which IL-8 production was measured in supernatant. Results are shown as the summary of all experiments (A) or pairwise (B), with each symbol representing an individual patient serum tested. (C) Inhibition of IL-8 production by mast cells in response to plate-bound ACPA, plate-bound IgG or lipopolysaccharide (LPS) by anti-CD32 antibodies or isotype control, indicated as a percentage of total IL-8 production in stimulated mast cells without blocking antibodies. (D) Mast cells were stimulated on CCP-coated plates incubated with the serum of ACPA+RF− patients. Results are shown as the summary (mean±SEM) of three independent mast cell donors using the serum of two different patients. (E) Mast cells were stimulated on CCP-coated plates incubated with ACPA IgG monoclonal antibody. Results are shown as the summary (mean±SEM) of four independent mast cell donors. (F) IL-8 production by mast cells in response to endogenous TLR ligand HSP70. (G) Inhibition of IL-8 production by mast cells in response to HSP70, by anti-TLR-2 or anti-TLR-4 antibodies, indicated as a percentage of total IL-8 production in stimulated mast cells without blocking antibodies. (H,I) IL-8 production by mast cells in response to combined triggering using plate-bound ACPA and HSP70 or their separate stimulations. Results are shown as the summary of all independent (n=5) experiments (H) or pairwise (I), with each symbol representing an individual patient serum tested. (I) IL-8 production after combined stimulation with HSP70 and ACPA (closed symbols) compared to the sum of IL-8 following separate stimulation with HSP70 and ACPA (open symbols). Asterisks indicate significant (p<0.05) differences between the indicated conditions, using paired samples t test. RF, rheumatoid factor.
Subsequently, we evaluated the activation of mast cells by the endogenous TLR ligand HSP70 (figure 6F). To confirm that activation with HSP70 was TLR dependent, mast cells were pre-incubated with blocking antibodies to TLR-2 and -4. As shown in figure 6G, activation of mast cells by HSP70 was inhibited by blocking TLR-4.
Combined activation of mast cells by ACPA immune complexes and HSP70 led to considerable increases in IL-8 production as compared to the single stimulus (figure 6H). This increase was significantly higher than the cumulative production of the separate stimuli when ACPA-positive serum was used, whereas no additional IL-8 production was observed when ACPA-negative serum was combined with HSP70 (figure 6I). Together, these results show enhanced cytokine production by mast cells when exposed to ACPA immune complexes and endogenous TLR ligands.
Discussion
In this study, we show that human mast cells can be activated by TLR triggering as well as FcγR ligation. Only limited information is available on the activation of human mast cells via TLR and FcγR. Here, we have extensively analysed mast cell responses towards ligands for all TLR ligands and found different patterns of cytokine production depending on the TLR that was triggered. Such different cytokine production profiles in response to TLR triggering have previously been reported for dendritic cells and basophils, and are thought to contribute different types of immune responses to different pathogens.31 ,32 Our results therefore indicate that different cytokines produced by mast cells may further promote these differences.
In addition to the response to pathogen-derived TLR ligands, we show that mast cells also respond to endogenous TLR ligands which are present in RA synovium. Endogenous ligands in synovium have been postulated to induce a positive feedback loop leading to chronic immune activation.13 ,33 Activation of TLR may be important for several chronic rheumatic diseases, including RA and spondyloarthritis (SpA).34 Since mast cells are relatively abundant in RA and SpA synovium,7 ,8 ,35 they may represent an important cell type involved in TLR-driven chronic inflammation.
Besides activation of mast cells via TLR ligands, we show for the first time that human mast cells can be triggered by ACPA immune complexes in a citrulline-dependent manner. Human mast cells were activated via IgG-ACPA, mediated by FcγRIIA.36–38 FcγRI was not expressed by synovial mast cells in the majority of patients, which is also in line with the absence of FcγRI expression by mast cells in other tissues.34–36 ,38 As ACPA immune complexes activated mast cells mainly via FcγRIIA and as this receptor is expressed by synovial mast cells from all patients analysed, we consider it most likely that this receptor is responsible for mast cell activation by ACPA immune complexes.
A clear limitation of our study is that it is not feasible to perform functional assays using mast cells isolated from synovium. However, although we cannot show that synovial mast cells respond in the same way as cultured mast cells, the cultured mast cells generally resemble the phenotype of synovial mast cells in their granule composition and in their expression of FcγR.
Importantly, we showed that combination of IgG- and TLR-mediated activation greatly enhanced cytokine production by human mast cells. Synergy between TLR and Fc receptor triggering has been previously described for basophils and dendritic cells,32 ,39 and likely represents a physiological function of the immune system to mount an enhanced response when antibodies are produced after the first encounter with a pathogen.
Synergy in the production of chemokines may dictate the type and extent of immune cells which are attracted to the site of inflammation,40 and may therefore also have detrimental effects in the context of autoimmunity. For example, IL-8 is increased in synovial fluid of RA patients and was shown to contribute to 30–60% of neutrophil chemoattraction by synovial fluid.41 Therefore, synergistic IL-8 production by mast cells in response to ACPA immune complexes and endogenous TLR ligands may lead to increased neutrophil recruitment, and thus contribute to inflammatory processes in RA.
In conclusion, we found that human mast cells functionally express TLRs and FcγRIIA and that triggering of TLR via endogenous TLR ligands can synergise with IgG-ACPA-mediated activation, greatly increasing cytokine production. These results suggest a role for combined TLR and FcγRIIA triggering of mast cells in the pathogenesis of RA.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Files in this Data Supplement:
- Data supplement 1 - Online figures
- Data supplement 2 - Online table 1
Footnotes
Handling editor Tore K Kvien
Funding This work was supported by the Dutch Arthritis Foundation, the Dutch Organization for Scientific Research (Vici grant), the Research Foundation Sole Mio, the Leiden Research Foundation (STROL), the Centre for Medical Systems Biology (CMSB) within the framework of the Netherlands Genomics Initiative (NGI), the IMI JU funded project BeTheCure, contract no. 115142-2, and the European Union (Seventh Framework Programme integrated project Masterswitch; grant no. 223404).
Competing interests None.
Ethics approval Leiden University Medical Center human ethics committee approved this study.
Provenance and peer review Not commissioned; externally peer reviewed.