Article Text

Abstract
Objective Primary Sjögren's syndrome (pSS) is characterised by chronic hyperactivation of B lymphocytes. Salivary gland epithelial cells (SGECs) could play a role in promoting B-lymphocyte activation within the target tissue. We aimed to study the interactions between SGECs from patients with pSS or controls and B lymphocytes.
Methods Patients had pSS according to 2016 European League Against Rheumatism/American College of Rheumatology criteria. Gene expression analysis of SGECs and B lymphocytes from pSS and controls isolated from salivary gland biopsies and blood was performed by RNA-seq. SGECs from pSS and controls were cocultured with B-lymphocytes sorted from healthy donor blood and were stimulated. Transwell and inhibition experiments were performed.
Results Gene expression analysis of SGECs identified an upregulation of interferon signalling pathway and genes involved in immune responses (HLA-DRA, IL-7 and B-cell activating factor receptor) in pSS. Activation genes CD40 and CD48 were upregulated in salivary gland sorted B lymphocytes from patients with pSS. SGECs induced an increase in B-lymphocyte survival, which was higher for SGECs from patients with pSS than controls. Moreover, when stimulated with poly(I:C), SGECs from patients with pSS induced higher activation of B-lymphocytes than those from controls. This effect depended on soluble factors. Inhibition with anti-B-cell activating factor, anti-A proliferation-inducing ligand, anti-interleukin-6-R antibodies, JAK1/3 inhibitor or hydroxychloroquine had no effect, conversely to leflunomide, Bruton's tyrosine kinase (BTK) or phosphatidyl-inositol 3-kinase (PI3K) inhibitors.
Conclusions SGECs from patients with pSS had better ability than those from controls to induce survival and activation of B lymphocytes. Targeting a single cytokine did not inhibit this effect, whereas leflunomide, BTK or PI3K inhibitors partially decreased B-lymphocyte viability in this model. This gives indications for future therapeutic options in pSS.
- Sjøgren's syndrome
- B cells
- autoimmune diseases
Statistics from Altmetric.com
Key messages
What is already known about this subject?
Primary Sjögren’s syndrome (pSS) is characterised by a chronic hyperactivation of B lymphocytes, and salivary gland epithelial cells (SGECs) could be involved in pSS pathophysiology.
What does this study add?
The RNA-seq analysis of SGECs sorted from salivary glands showed upregulated interferon (IFN) signalling pathway, HLA-DR, IL-7 and BAFFR. The RNA-seq analysis of B lymphocytes sorted from salivary glands showed upregulated CD40 and CD48.
In a coculture model, this study showed that survival and activation of B lymphocytes were higher with SGECs from patients with pSS than controls.
Interactions between SGECs and B lymphocytes were driven mainly by secretion of soluble factors.
How might this impact on clinical practice or future developments?
Targeting a single cytokinewas not sufficient to inhibit this effect, but leflunomide, BTK or PI3K inhibitors were successful, which suggests future therapeutic options in pSS.
Introduction
Primary Sjögren's syndrome (pSS) is a systemic autoimmune disorder characterised by a lymphocytic infiltrate of exocrine glands. Patients present with sicca, fatigue and pain, and 30%–40% of patients present with systemic manifestations, including involvement of skin, joints, neurological system, lung or kidney.1 B-lymphocyte hyperactivation is a hallmark of the disease,2 as illustrated by the presence of serum autoantibodies, hypergammaglobulinaemia, increased levels of free light chains and increased risk of B-cell lymphoma. The periepithelial tropism of the lymphocytic infiltration is specific to pSS and justifies the term autoimmune epithelitis.3
Several lines of evidence support that salivary gland epithelial cells (SGECs) not only are the target of autoimmunity in patients with pSS but also may play a role in the pathology. They could act as non-professional antigen-presenting cells by expressing costimulatory molecules such as CD80 and CD86.4 In salivary glands from patients with pSS, Cytometry by time-of-flight (CyTOF) immunophenotyping showed a significant upregulation of human leukocyte antigen (HLA)-DR in SGECs associated with disease activity.5 SGECs could be involved in local B-lymphocyte activation as they can secrete B-cell activating factor (BAFF).6 Of note, plasma cells are detected in salivary gland tissue in close contact with CXCL12-expressing ductal and acinar epithelia, which suggests potential interactions between these cells.7
We hypothesised that SGECs could play a supportive role in B-lymphocyte survival and activation. We first focused on gene expression analysis of purified cellular populations sorted from salivary gland biopsies, then developed an in vitro coculture model, including SGECs and B lymphocytes to characterise the cross-talk between both.
Methods
Patients
Minor salivary gland (MSG) biopsies were obtained from consecutive patients referred for suspected pSS to the rheumatology department of Bicêtre Hospital, a tertiary reference centre for systemic autoimmune diseases. pSS was defined according to the 2016 American College of Rheumatology/European League Against Rheumatism criteria,8 and controls were patients presenting sicca symptoms without anti-Ro/SSA (Sjögren's syndrome related Antigen A) or anti-La/SSB antibodies and with normal or subnormal MSGs (ie, focus score of <1). MSG biopsies were used for transcriptomic analysis of sorted cells and coculture experiments. Peripheral blood mononuclear cells (PBMCs) used for transcriptomic analysis were isolated from whole blood of patients with pSS and healthy blood donors. B lymphocytes sorted from blood of healthy donors were used for the coculture experiments.
A total of 42 patients with pSS and 62 controls were included in the study, among whom 26 patients with pSS and 27 controls had an MSG biopsy. The characteristics of the patients and controls are presented in table 1. The experimental design showing the number of patients included in each experiment is shown in online supplementary figure S1A.
Supplemental material
Characteristics of the patients with pSS and controls
Patients or the public were not involved in the design, conduct, reporting or dissemination plans of our research.
Methods of RNA-sequencing, primary cultures of SGECs9 and cocultures of SGECs and B lymphocytes, FACS analysis and assays are described in the online supplementary text.
Statistical methods
Statistical analysis of RNA-seq
Statistical analyses were performed using the DESeq2 package. Comparisons were performed between patients with pSS and controls in each cell subtype based on a non-parametric t-test. Cells sorted from biopsies and from blood in four patients with pSS (with matched samples available) underwent paired comparison by non-parametric paired t-test. A p value of <0.05 was considered statistically significant to identify differentially expressed genes.
Statistical analysis of coculture experiments
The percentage of surviving B lymphocytes was calculated for each culture condition as the difference between the percentage proportion of alive B lymphocytes cocultured with SGECs and the percentage of alive B lymphocytes cultured alone. The same strategy was used to assess the percentage of activated B lymphocytes (CD38+ or CD48+ B-lymphocyte percentages and mean fluorescence intensity (MFI)). Statistical analyses were performed with Prism software using Mann-Whitney (unpaired data) and Wilcoxon (paired data) tests.
Results
SGECs from patients with pSS overexpress interferon (IFN)-induced genes, HLA-DRA, interleukin (IL)-7 and B-cell activating factor receptor (BAFFR)
Differential gene expression analysis of SGECs from five patients with pSS vs four controls revealed 494 genes differentially expressed, 280 upregulated and 214 downregulated. IFI6, VGLL2 and ZNF879 were the most significantly upregulated genes, and CCL22 was one of the most significantly downregulated genes (−log10 (p-value) > 4) (figure 1A). The identified transcriptional differences were associated with disease status as indicated by the distinct clustering of pSS and control samples in the multidimensional scaling (MDS) plot shown in figure 1B. Functional enrichment analysis of differentially expressed genes highlighted an over-representation of the IFN signalling pathway, B-cell development pathway and IL-7 signalling pathway (−log10(p-value)=3.52, 2.89 and 2.51, respectively) (online supplementary table S3). Several IFN-induced genes were upregulated, including IFI6, OAS2, STAT1 and BST2 (bone marrow stromal cell antigen 2) (figure 1C). Among the 280 upregulated genes in pSS SGECs, 18 were IFN-induced genes according to the Interferome database; 8 were induced by types I and II; 9 were induced by types I, II and III; and 1 was induced by type I only. We also observed an upregulation of HLA-DRA expression, coding for the alpha chain of the DR major histocompatibility complex, and IL-7 expression in SGECs from patients with pSS versus controls (p=0.04, p=0.02, respectively) (figure 1C and online supplementary table S4). Of note, for all these genes, the highest gene expression values were obtained in anti-SSA positive patients who also had the highest salivary gland infiltration. These results were confirmed by reverse transcription (RT)-PCR, statistically significant for BST2 and IFI6 and with a trend for HLA-DRA, OAS2, STAT1 and IL-7 (online supplementary figure S3). Lastly, an upregulation of BAFFR expression was observed in SGECs from patients with pSS versus controls (p=0.01) with RNA-seq (figure 1C) and confirmed by flow cytometry of unstimulated SGECs, with no change after poly(I:C) stimulation (figure 1D,E).

SGECs sorted from salivary gland biopsies express IFN-induced genes, HLA-DRA, BAFFR, Protein Kinase R (PKR) and IRF7. (A) Volcano plot of differentially expressed genes in sorted SGECs from patients with pSS (n=5) versus CTRLs (n=4). The horizontal red dashed line indicates the cut-off for significance at p<0.05 and the vertical lines for fold change in expressions ≥1.5 and ≤−1.5 (B) Multidimensional scaling representation of the similarities between transcriptomic profiles. Each dot in the representation corresponds to a transcriptomic profile, and the distances between the dots are proportional to the transcriptomic distances computed based on the list of 494 differentially expressed genes.(C) Normalised counts of genes expressed by SGECs in healthy controls (HC) and patients with pSS. Results of RNA-seq of SGECs sorted cells: IFN-stimulated genes (IFI6, OAS2, STAT1 and BST2), HLA-DRA, IL-7, BAFFR, PKR and IRF7. P values were determined by using DESeq2 software. (D) Flow cytometry example of BAFFR expression in unstimulated alive SGECs and isotype CTRL from a patient with pSS. (E) Expression of BAFFR evaluated by flow cytometry on alive SGECs with or without poly(I:C) stimulation. Statistical analysis was performed using the Mann-Whitney test. SGECs from CTRLs are in green; SGECs from patients with pSS are in orange. *, corresponds to a p value of ≤0.05. BAFFR, B-cell activating factor receptor; CTRL, control; IFN, interferon; IL, interleukin; ns, not significant; pSS, primary Sjögren’s syndrome; SGEC, salivary gland epithelial cell; SSC-A, side scatter-A.
B-lymphocyte activation is observed at the transcriptomic level in salivary gland biopsies and in blood
When comparing gene expression in sorted B lymphocytes from salivary glands of patients with pSS and controls, we detected 620 differentially expressed genes, 397 upregulated and 223 downregulated. The most significantly upregulated genes were IGHG1 and HADH, and the most significantly downregulated genes were MT-RNR2-L12, MARCO, S100A9, ELF4 and ITGA1 (−log10 (p-value) >5) (figure 2A). The identified differentially expressed genes were associated with the disease status as indicated by the distinct clustering of pSS and control samples in the MDS plot (figure 2B). Several genes involved in B-lymphocyte activation and differentiation, such as CD40, CD48 and CD22 (p=0.02, p=0.01 and p=0.048, respectively), were upregulated in patients with pSS versus controls (figure 2C). CD40 and CD48, but not CD22, upregulation was confirmed by RT-PCR (online supplementary figure S3). The IFN-induced gene, OAS2 and IFN-γ receptor 2 were also upregulated in B lymphocytes from patients with pSS versus controls (p=0.04 and p=0.03, respectively) but were not confirmed by RT-PCR. The absence of RT-PCR confirmation may be due to a lack of power.

B lymphocytes sorted from salivary gland biopsies from pSS overexpress activation and differentiation genes as compared with CTRLs. (A) Volcano plot of differentially expressed genes in sorted B lymphocytes from patients with pSS (n=9) versus CTRLs (n=4). The horizontal red line indicates the cut-off for significance at p<0.05 and the vertical lines indicate fold changes at ≥1.5 and ≤−1.5. (B) Multidimensional scaling representation of similarities between transcriptomic profiles. Each dot corresponds to a transcriptomic profile, and distances between the dots are proportional to the transcriptomic distances computed based on the 620 differentially expressed genes.(C) Normalised counts of CD40, CD22, CD48, OAS2 and IFNGR2 in B lymphocytes sorted from biopsies from controls (HC, healthy controls) and in patients with pSS. P values were determined by using DESeq2. CTRL, control; IFNGR2, IFN-γ receptor 2; pSS, primary Sjögren’s syndrome.
In sorted B lymphocytes from blood, 516 genes were differentially expressed between patients with pSS and controls, 359 upregulated and 157 downregulated. Functional enrichment pathway analysis highlighted an over-representation of EIF2 signalling and IFN signalling pathways, as well as a role for JAK signalling (online supplementary table S3). TLR7 and its downstream signalling molecule, IRF7, were upregulated in pSS; additionally, IL-6, involved in B lymphocyte growth, was upregulated (online supplementary table S4). The RT-PCR results showed a trend for a higher expression of TLR7, IRF7 and IL-6 in pSS compared with controls (online supplementary figure S3C).
The paired comparison between B lymphocytes from salivary glands and from blood in four patients with pSS revealed the upregulated genes CD138, a plasma cell marker; RANK; and IL-6. Moreover, CCR10, which codes for the receptor for CCL28, a mucosal homing chemokine, was upregulated in B lymphocytes sorted from biopsies versus blood (online supplementary table S4). These results were confirmed for RANK and IL-6 by RT-PCR (p=0.03 and p=0.03).
SGECs increase B-lymphocyte survival and activation
To analyse the potential interaction between SGECs and B lymphocytes, we performed 5 days of cocultures of SGECs from patients with pSS or controls with B lymphocytes from healthy donors with or without IFN-α, IFN-γ or poly(I:C) stimulation. We observed an increase of B-cell viability under all stimulation conditions (p<0.001) (figure 3A,B). The increase in B-lymphocyte survival was higher in the poly(I:C) than in the IFN-α or IFN-γ-stimulated cocultures (figure 3A and online supplementary figure S4). Conversely, poly(I:C) did not drive survival of B lymphocytes cultured alone (figure 3A,B). More importantly, survival of B lymphocytes was higher in coculture with SGECs from patients with pSS (stimulated or not with poly(I:C)) than in coculture with SGECs from controls (p<0.05) (figure 3C).

SGECs from patients with pSS increase B-lymphocyte survival and activation. (A) Representative dot plot of B-lymphocyte viability stained with live/dead (fixable viability dye) cultured alone and or cocultured with SGECs with or without poly(I:C). (B) Percentage of alive B lymphocytes on day 5, cultured alone or cocultured with SGECs with or without IFN-α, IFN-γ or poly(I:C). (C) Difference between the percentage of alive B lymphocytes cocultured with SGECs from patients with pSS and controls and the percentage of alive B lymphocytes cultured alone on day 5. (D) Representative dot plot of B lymphocytes cultured alone and cocultured with SGECs with or without poly(I:C) and stained with for CD38 and CD27. (E) Percentage of CD38+ B lymphocytes at day 5 in all conditions of culture and stimulation. (F) Difference between the percentage of CD38+ B lymphocytes in coculture with SGECs and the percentage of CD38+ B lymphocytes cultured alone at day 5. (G) MFI of CD38 in B lymphocytes cultured alone with or without poly(I:C) or B lymphocytes cocultured with SGECs with or without poly(I:C). (H) Difference between the MFI for CD38 for B lymphocytes cocultured with SGECs or cultured alone on day 5. *P≤0.05, **P≤0.01, ***P≤0.001. IFN, interferon; MFI, mean fluorescence intensity; pSS, primary Sjögren’s syndrome; SGEC, salivary gland epithelial cell.
Cocultures of SGECs with B lymphocytes also increased B-lymphocyte activation, as assessed by increased expression of CD38 on B lymphocytes under all stimulation conditions (figure 3D,E), with a difference between SGECs from patients and controls after poly(I:C) stimulation (figure 3F,H). Poly(I:C) had a minor impact on the percentage and the MFI of CD38 on B lymphocytes cultured alone (figure 3E,G).
The effect of SGECs from patients with pSS on B-lymphocyte viability and CD38 expression was increased by poly(I:C) but not IFN stimulation. Thus, we wondered whether Toll-like receptor 3 (TLR3) signalling might differ between patients and controls. However, on looking at the poly(I:C) signalling pathways in SGECs, we found no difference between pSS and controls in the expression of TLR3, TRAF3, IRF3, RIG1 and MDA5 (data not shown). However, PKR and IRF7 were upregulated but not significantly in SGECs from patients with pSS versus controls (figure 1C). Of note, no expression of TLR3 in B lymphocytes sorted from salivary gland biopsies and from blood was detected.
After coculture with SGECs, the proportion of CD27+ B lymphocytes was increased, and notably switched memory B lymphocytes defined as CD27+ IgD cells on coculture with poly(I:C) stimulation (online supplementary figure S5A,B). The coculture with SGECs did not achieve differentiation of B lymphocytes into antibody-secreting cells, as supported by the lack of increase in immunoglobulin (IgM and IgG) production in the coculture supernatants (online supplementary figure S6A,B).
To sum up, the coculture experiments demonstrated that SGECs from patients with pSS had a higher ability than SGECs from controls to increase survival and activation of B lymphocytes.
Interactions between SGECs and B lymphocytes require mostly soluble factors
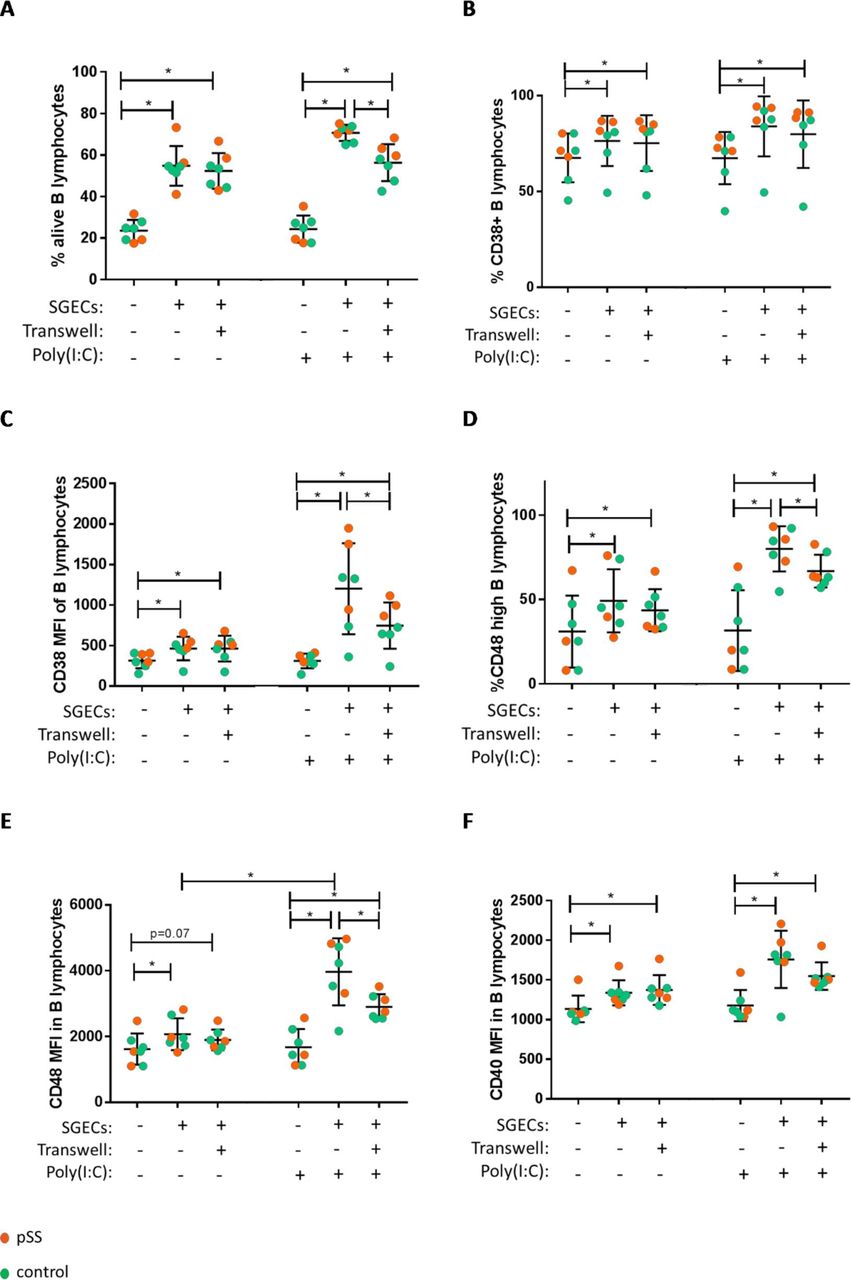
To determine whether the survival and activation of B lymphocytes required direct cell contact or not, we performed transwell experiments. In the absence of stimulation, the transwell separation did not change the percentage of surviving B lymphocytes cocultured with SGECs, which suggests that B-lymphocyte survival mostly depended on soluble factors (figure 4A). Similar results were obtained for B-lymphocyte activation as assessed by CD38, CD48 and CD40 markers (figure 4B–F). However, under poly(I:C) stimulation, part of the effect of SGECs on B-lymphocyte survival and activation was reduced with transwell separation, which suggests the additional role of cell contact in this specific condition (figure 4A,C–E).

Increases in B-lymphocyte viability and activation were mainly due to soluble factors. B lymphocytes were examined on day 5 with and without transwell, with and without poly(I:C), in cocultures with SGECs from patients with pSS and controls. (A) Percentage of alive B lymphocytes. (B) Percentage of CD38+ B lymphocytes. (C) CD38 MFI of B lymphocytes. (D) Percentage of CD48 high B lymphocytes. (E) CD48 MFI of B lymphocytes. (F) CD40 MFI of B lymphocytes. *P≤0.05. MFI, mean fluorescence intensity; pSS, primary Sjögren’s syndrome; SGEC, salivary gland epithelial cell.
Then, in order to determine which factors could play a role in the interactions between SGECs and B lymphocytes, we tested successively potential involved factors (including BAFF, A proliferation-inducing ligand (APRIL) and IL-6) and signalling pathways (including JAK, Bruton's tyrosine kinase (BTK), Phosphatidyl-inositol 3-kinase (PI3K) and pathways involved in B-lymphocyte activation and proliferation) by adding specific inhibitors in the cocultures. In spite of detection of numbers of cytokines in the supernatant of cocultures (figure 5A–D), especially in patients with pSS, none of the tested anticytokines (belimumab, anti-APRIL antibody and tocilizumab (anti-IL-6-R)) was effective for decreasing B-lymphocyte survival and activation by SGECs. Likewise, tofacitinib (JAK inhibitor) was unsuccessful (figure 5I,J). Several chemokines were detected in coculture supernatants, which suggests a potential role for homing factors secreted by SGECs supporting B-lymphocyte survival and activation. CXCL10, also known as IFN-γ-inducible protein 10, and CXCL13 were detected in SGECs cultures and coculture supernatants (figure 5E,G). At day 5, CXCL12 was detected at very low levels in the supernatant of only three pSS SGEC cocultures (figure 5F) but not in control SGECs cocultures. CXCL12 was detected in SGECs cultures supernatants at days 1 and 2 (online supplementary figure S7). Lastly, CCL28, another chemokine involved in mucosal homing, was detected in the coculture supernatant, with no difference between patients with pSS and controls (figure 5H).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Assessment of soluble factors in coculture supernatants and inhibition experiments. (A) Level of A proliferation inducing ligand (APRIL) in the supernatants of B lymphocytes cultured alone, SGECs from pSS and controls cultured alone and SGECs cocultured with B lymphocytes.(B) Dosage of APRIL in supernatants of SGECs cultured alone or cocultured with B lymphocytes (after subtraction of APRIL level in supernatants of B lymphocytes cultured alone). (C) Level of IL-6 in supernatants of B lymphocytes cultured alone, SGECs from pSS and controls cultured alone and SGECs cocultured with B lymphocytes. (D) Level of IL-6 in supernatants of SGECs cultured alone or cocultured with B lymphocytes (after subtraction of IL-6 level in supernatants of B lymphocytes cultured alone). (E–H) Level of CXCL10 (E), CXCL12 (F), CXCL13 (G) and CCL28 (H) in supernatants of B lymphocytes cultured alone, SGECs from pSS and controls cultured alone and SGECs cocultured with B lymphocytes. Dashed line represents the lower limit of detection of the assay. (I,J): Viability of B lymphocytes cocultured with SGECs and different inhibitors relative to the condition without the inhibitor and without poly(I:C) (I) and with poly(I:C) (J). (K) Effect of leflunomide, ibrutinib (Bruton's tyrosine kinase (BTK) inhibitor) and LY294002 (Phosphatidyl-inositol 3-kinase (PI3K) inhibitor) on CD38 MFI of B lymphocytes cultured alone and in coculture with SGECs. *P≤0.05, **P≤0.01, ***P≤0.001. IL, interleukin; pSS, primary Sjögren’s syndrome; SGEC, salivary gland epithelial cell.
Given these results, we tried to target multiple immunological pathways at the same time by using BTK and PI3K inhibitors. BTK was targeted by adding ibrutinib, a BTK inhibitor, in the cocultures. Ibrutinib decreased B-lymphocyte viability in coculture (p<0.05), but this effect was lost in the presence of poly(I:C) (figure 5I,J). LY294002, a PI3K inhibitor, decreased B-lymphocyte viability in coculture (p<0.05) (figure 5I). This effect was maintained but not significantly with poly(I:C) stimulation (figure 5J). LY294002 could also decrease cocultured B-lymphocyte activation, as assessed by the decrease in CD38 MFI (figure 5K). Of note, there was no effect of the leflunomide, the BTK inhibitor and the PI3K inhibitor on SGEC viability as compared with the control condition (DMSO). LY294002, but not leflunomide or ibrutinib, also affected B-lymphocyte viability when B lymphocytes were cultured alone (online supplementary figure S8).
Given some recent clinical results,10 we also looked at the effect of the association of leflunomide and hydroxychloroquine. Leflunomide decreased B-lymphocyte viability in coculture, but this effect was lost in the presence of poly(I:C) (figure 5I,J). Hydroxychloroquine alone or combined with leflunomide had no effect on cocultured B-lymphocyte viability (figure 5I).
Discussion
Transcriptome analysis of sorted cells from salivary gland biopsies showed an increased IFN signature in SGECs and a higher activation of tissue and circulating B lymphocytes in patients with pSS than controls. Using a coculture model, SGECs from patients with pSS were more efficient to promote survival and activation of B lymphocytes than SGECs from controls. These effects were mainly mediated by soluble factors. Targeting a single cytokine (BAFF, APRIL, IL-6 and JAK1/3 pathway) did not inhibit this effect, whereas, leflunomide, BTK or PI3K inhibitors partially decreased B-lymphocyte viability induced by SGECs. A schematic representation of this hypothesis is presented in online supplementary figure S9. Important roles for B lymphocytes, SGECs and IFN have been suggested by previous studies in pSS pathophysiology. This work brings new elements to better characterise the interactions between these actors. Moreover, the results we obtained in this model could have therapeutic consequences by suggesting that directly targeting B-lymphocyte activation pathways would be more efficient than targeting a single cytokine. However, our model with only two cellular actors is oversimplified, and we cannot eliminate that, in vivo, anticytokine therapy might have an indirect action on the microenvironment.
RNA-seq analysis of sorted SGECs demonstrated an IFN signature within these cells. The IFN signature is well known in PBMCs and salivary gland biopsies of patients with pSS11 12; however, this is the first time that this IFN signature has been demonstrated in sorted SGECs. The chronic expression of endogenous retroviral (ERV) sequences could explain the over-representation of the IFN signalling pathway in SGECs from patients with pSS.13 Overexpression of BST2, a transmembrane protein that inhibits viral particles budding from infected cells, could suggest an overexpression of some ERVs. A recent work highlighted increased mRNA and protein levels of BST2 in salivary glands from patients with pSS.14 However, in our study, BST2 overexpression was only the consequence of IFN overexpression since we did not find a differential expression of coding ERV between MSG from patients with pSS and controls (data not shown).
SGECs could act as non-professional presenting cells by expressing HLA-DR but also adhesion and costimulatory molecules.15 We observed an upregulation of HLA-DRA, which codes for one subunit of HLA-DR. This observation agrees with previously published works showing an aberrant expression of HLA-DR by SGECs,16 which can be induced by a type II IFN stimulation.17
Type I and II IFNs are also able to induce BAFF production by SGECs.6 BAFF could represent the link between the innate and adaptive immunity in pSS. BAFFR is the most specific receptor of BAFF and is mainly present in B lymphocytes. By using RT-PCR and immunofluorescence, Lahiri et al 18 showed that BAFFR was expressed by SGECs and that this receptor was involved in SGEC survival owing to an autocrine effect of BAFF. We showed a differential expression of BAFFR on SGECs between pSS and controls. Thus, the BAFF cytokine could play a dual role, having a direct effect on B-lymphocyte survival and also supporting SGEC survival, which in turn may have a feeder effect on B lymphocytes.
B lymphocytes sorted from salivary glands from patients with pSS overexpressed activating genes, as expected based on the pSS pathophysiology. Of note, the coculture between SGECs and B lymphocytes increased the expression of activation markers, such as CD38, CD48 and CD40, as compared with B lymphocytes cultured alone. CD40, a costimulatory pathway receptor essential for germinal centre reactions, seems of particular interest.19–21 Moreover, a phase II therapeutic trial of an anti-CD40 antibody recently yielded positive results.22 In our study, CD48, which may enhance the CD40-mediated activating signal for B lymphocytes,23 was upregulated in pSS versus controls. This result was congruent with a work describing soluble CD48 as a biomarker of pSS, which was correlated with the biological domain of the ESSDAI.24
B lymphocytes sorted from blood overexpressed TLR7 and the downstream signalling molecule IRF7. Interestingly, the upregulation of IRF7 was confirmed by Sun et al,25 who recently performed RNA-seq analysis of B lymphocytes sorted from blood comparing three patients with pSS to three controls (log2 fold change (FC)=1.39 and p=8.59x 10–7) (online supplementary figure S10). Moreover, Imgenberg-Kreuz et al 26 showed in a RNA-seq analysis of B-lymphocytes sorted from blood an upregulation of TLR7 as well as IRF7 in pSS compared with controls (FC=2.8, p=5.09x10−5 and FC=2.8, p=5.09x10−5, respectively).
Previously, coculture experimental models with SGECs used allogenic PBMCs and failed because of allogenic reactions.17 In our coculture model, we used sorted allogenic B lymphocytes, avoiding the T-lymphocyte allogenic reaction. Moreover, the effect of SGECs on B-lymphocyte survival and activation was maintained even with transwell separation that would have prevented antigen presentation. Coculture with a human salivary gland cell line and tonsillar B lymphocytes revealed increased apoptosis in epithelial cells.27 This model using a cancer cell line was more artificial than primary cultured SGECs from patients or controls. One limitation of our model is that we used B lymphocytes sorted from blood, which might have different characteristics than B lymphocytes sorted from biopsies. However, the use of B lymphocytes sorted from biopsies would have been not technically feasible due to the low number of B-lymphocytes present in salivary gland biopsies.
In our coculture model, the ability of SGECs from patients with pSS to increase B-lymphocyte survival was increased by poly(I:C) rather than IFN-α or IFN-γ stimulation. The transcriptomic analysis showed a higher expression of PKR and IRF7, although not significant, in SGECs from patients with pSS than controls, whereas the expression of other downstream signalling factors of TLR3 were similar. Thus, poly(I:C) stimulation could mimic a viral trigger and signal through several pathways inducing a positive feedback loop.
We hypothesised that SGECs could provide a favourable environment enhancing B-lymphocyte survival and activation. We observed an increase in memory B-lymphocyte differentiation in cocultures; however, the SGECs did not achieve differentiation of B lymphocytes in antibody-secreting cells. Thus, this coculture model remains partial, and other actors such as CD40L, IL-2, IL-4 and IL-21, as shown by McCarthy, using single-cell Nojima cultures could be required.28 T-follicular helper lymphocytes might also be required to obtain differentiated plasma cells.
By analogy with bone-marrow plasma cells niches, in which several survival factors, including BAFF, APRIL, IL-6 and CXCL12, are required,29 we assessed BAFF, APRIL, IL-6, CXCL10, CXCL12, CXCL13 and CCL28 in coculture supernatants. All were present in coculture supernatants and could participate in the survival and activation of B lymphocytes, CXCL12 at very low level and BAFF only after poly(I:C) stimulation, which agrees with our previous studies.6 30 Of note, CXCL12 was detected in the SGEC supernatants at days 1 and 2 of culture, but not at day 5 (online supplementary figure S7). This chemokine could be involved in B-lymphocyte recruitment and organisation of the tertiary lymphoid structures in the salivary glands.31 We did not use any specific inhibitor of the CXCL12 signalling, but this could be of interest. Our experiments with inhibitory molecules showed that a single pathway inhibition (anti-BAFF, anti-APRIL and anti-IL-6-R antibody) was not sufficient. Even tofacitinib, which targets numbers of proinflammatory cytokines, was not successful. Leflunomide, a pyrimidine synthesis inhibitor that inhibits B-lymphocyte proliferation and cell cycle progression,10 32 as well as PI3K and BTK inhibitors (targeting at the same time CXCL13, BAFF and IL-6 signalling33) decreased B-lymphocyte viability in cocultures. Lastly, PI3K was the only inhibitor to maintain an inhibitory effect under poly(I:C) stimulation, which suggests that in a local IFN-rich environment, such as pSS salivary glands, PI3K inhibition might be the most efficient option.
In conclusion, the analysis of SGECs and B lymphocytes sorted from salivary glands from patients with pSS showed an upregulation of several genes and pathways that could be involved in their interactions. Our coculture model showed that SGECs from patients with pSS had better ability than controls to stimulate survival and activation of B lymphocytes. This effect was mediated mostly by soluble factors. Targeting a single cytokine did not inhibit this effect, whereas, leflunomide, BTK or PI3K inhibitors partially decreased B-lymphocyte viability in this model. These findings may give indications for the choice of future therapeutic options in pSS.
Acknowledgments
We thank C Le Pajolec, Assistance Publique-Hôpitaux de Paris, Hôpitaux Universitaires Paris-Sud, Le Kremlin-Bicêtre, France; E. Berge, Rheumatology, Université Paris Sud, Le Kremlin Bicêtre, France, who participated in the recruitment of patients, and V Libri, Center for Translation Research, Institut Pasteur, Paris, France, for her expertise and help with the Biomark experiments. We thank Odile Heidmann, Kevin De Azevedo and Thierry Heidmann, Unité des Rétrovirus Endogènes et Eléments Rétroïdes des Eukaryotes Supérieurs, UMR 8122 CNRS, Institut Gustave Roussy, Villejuif, France, for their expertise and work on the endogenous retroviral sequences analysis. We thank Pascal Schneider, Department of Biochemistry, University of Lausanne, Switzerland, for providing a human APRIL blocking antibody.
References
Footnotes
GN and XM are joint senior authors.
Handling editor Josef S Smolen
GN and XM contributed equally.
Contributors ER participated in designing the research studies, conducting experiments, acquiring data, analysing data and writing the manuscript. JP participated in designing the research studies, conducting experiments, acquiring data and analysing data. NT participated in the analysis of the data and in writing the manuscript. SB participated in designing the research studies, conducting experiments and acquiring data. AP, BL and AD participated in designing the research studies, conducting experiments and acquiring data. HC, AT and NA participated in conducting experiments and acquiring data. BJ participated in acquiring data, analysing data and writing the manuscript. MM participated in designing the research studies, acquiring data, analysing data and writing the manuscript. GN and XM participated in designing the research studies, analysing data and writing the manuscript.
Funding Support was obtained from the Labex in Research on Medication and Therapeutic Innovation (ANR10), the Fondation pour la Recherche Médicale DEQ20150934719, an unrestricted grant from Biogen to Université Paris-Sud (UPSud/SAIC N 97731) and the Innovative Medicines Initiative 2 Joint Undertaking (JU) (NECESSITY grant agreement number 806975). This JU received support from the European Union’s Horizon 2020 research and innovation programme and EFPIA. ER is the recipient of a PhD fellowship from Arthritis Fondation Courtin, Arthritis R&D (CIFRE 2016/1406).
Competing interests AT, NA and MM are employed by Biogen. XM received an honorarium for consultancy advice on Sjögren’s syndrome from BMS, GSK, Novartis and Servier and a research grant from Servier. The rest of the authors declare that they have no relevant conflicts of interest.
Patient and public involvement Patients and/or the public were not involved in the design, or conduct, or reporting, or dissemination plans of this research.
Patient consent for publication Not required.
Ethics approval This study received approval from the local ethics committee, and informed consent was obtained from all participants.
Provenance and peer review Not commissioned; externally peer reviewed.
Data availability statement Data are available on reasonable request. All data relevant to the study are included in the article or uploaded as online supplementary information. Additional data are available on reasonable request.