Article Text

Abstract
Introduction The second generation anticycliccitrullinated peptide (anti-CCP2) assay detects the majority but not all anticitrullinated protein/peptide antibodies (ACPA). Anti-CCP2-positive rheumatoid arthritis (RA) is associated with HLA-DRB1* shared epitope (SE) alleles and smoking. Using a multiplex assay to detect multiple specific ACPA, we have investigated the fine specificity of individual ACPA responses and the biological impact of additional ACPA reactivity among anti-CCP2-negative patients.
Methods We investigated 2825 patients with RA and 551 healthy controls with full data on anti-CCP2, HLA-DRB1* alleles and smoking history concerning reactivity against 16 citrullinated peptides and arginine control peptides with a multiplex array.
Results The prevalence of the 16 ACPA specificities ranged from 9% to 58%. When reactivity to arginine peptides was subtracted, the mean diagnostic sensitivity increased by 3.2% with maintained 98% specificity. Of the anti-CCP2-negative patients, 16% were found to be ACPA positive. All ACPA specificities associated with SE, and all but one with smoking. Correction for arginine reactivity also conveyed a stronger association with SE for 13/16 peptides. Importantly, when all ACPA specificities were analysed together, SE and smoking associated with RA in synergy among ACPA positive, but not among ACPA-negative subjects also in the anti-CCP2-negative subset.
Conclusions Multiplexing detects an enlarged group of ACPA-positive but anti-CCP2-negative patients with genetic and environmental attributes previously assigned to anti-CCP2-positive patients. The individual correction for arginine peptide reactivity confers both higher diagnostic sensitivity and stronger association to SE than gross ACPA measurement.
- ant-ccp
- autoantibodies
- rheumatoid arthritis
Statistics from Altmetric.com
Introduction
The first publication on the highly specific anticitrullinated protein/peptide antibodies (ACPA) already showed that there was vast epitope heterogeneity between individual patients with rheumatoid arthritis (RA).1 The anticyclic citrullinated peptide 2 (anti-CCP2) assay includes a mixture of peptides aiming at optimising diagnostic sensitivity while keeping the diagnostic specificity high.2 This test and other general ACPA tests have allowed the subcategorisation of patients with RA with respect to ACPA status.3 4
Since the discovery of ACPA, there has been an extensive search for citrullinated autoantigens inside and outside rheumatic joints with an aim to define structures of importance for the initiation and progression of ACPA responses in patients with RA. The list of autoantigens encompasses epitopes on, for example, filaggrin,5–8 fibrin/fibrinogen,9–12 vimentin,13–15 alpha-enolase16 and collagen type II.17 18 Such compartmentalisation of the ACPA response has proven useful. In 2009, Madhi et al showed that the smoking–HLA-DRB1* shared epitope (SE) association previously attributed to anti-CCP2-positive RA patients was mostly confined to patients double positive for antibodies against CCP2 and CEP-1, the immunodominant citrullinated alpha-enolase epitope, whereas patients single positive for anti-CCP2 showed a much lower degree of association.16 19 Such studies have since been repeated with larger panels of specific ACPA reactivities including citrullinated vimentin peptides and being used for the immunological subsetting of RA in the context of genes such as SE and PTPN22, as well as smoking.20 21
To facilitate such subsetting studies, we have developed a peptide microarray for the parallel detection of autoantibodies against multiple citrullinated peptides. A proof-of-concept study was published in 2012, where we investigated reactivity against 12 different citrullinated peptides in 927 patients with RA and 461 healthy controls from the Epidemiological Investigations in Rheumatoid Arthritis (EIRA) case–control study.22 We described varying sensitivity at the same predefined high specificity (98%) for individual ACPA, with some of them approaching but never surpassing the anti-CCP2 sensitivity. Although the majority of ACPA peptide-positive patients were also anti-CCP2 positive, there were a considerable number of ACPA peptide-positive patients also in the anti-CCP2 negative subset.
A number of studies have described non-specific anti-CCP2 responses in, for example, tuberculosis,23 24 hepatitis C,25 26 autoimmune hepatitis27 and Leishmania donovani infection.28 Such non-specific responses have been demonstrated by comparing the ELISA reactivity against the CCP2 with reactivity against identical plates coated with the parallel native peptides containing the original arginine residues instead of citrulline or only blocked wells. Due to conventional assay configuration, such individual controls are almost never performed in routine ACPA testing in clinical laboratories. The microarray format, however, easily allows simultaneous evaluation of reactivity against citrullinated peptides and their arginine-containing counterparts.22
In the present study, we have investigated the biological significance of additional ACPA reactivity among anti-CCP2-negative individuals, and we have related each individual ACPA response to reactivity with the analogue arginine-containing peptide.
Patients and methods
Subjects
Patients and controls from the EIRA case-control study were included, for the RA patients within 12 months after first symptoms. Patients were aged between 18 and 70 years and diagnosed according to the 1987 American College of Rheumatology (ACR) criteria by a rheumatologist, and sampling was performed at the first visit before disease-modifying anti-rheumatic drug (DMARD) treatment.29 Controls were matched for age, sex and residential area. Data on smoking habits were retrieved by questionnaire at inclusion, and patients and controls were genotyped for HLA-DRB1* and grouped in relation to SE alleles. More information about EIRA can be found elsewhere.3 30
In total 2934 patients and 624 controls were investigated. Due to the propensities of certain sera (eg, direct binding to all peptides containing streptavidin) and high background reactivity to the microarray surface, 118 samples (74 patients and 44 controls) were excluded. Two patients and two controls lacked anti-CCP2 data, 24 patients and 13 controls lacked information about HLA-DRB1* alleles and smoking information was missing for 24 patients and 13 controls. A total of 109 patients and 73 controls were excluded, leaving 2825 patients and 551 controls for the final analysis.
Serum samples were drawn at the time of inclusion in EIRA and thereafter stored frozen at −70°C. All subjects had given informed consent, and ethical approval was granted by the Karolinska Institutet ethics committee.
ACPA microarray
ACPA reactivity was measured using a custom-made microarray based on the ImmunoCAP ISAC system (Phadia AB, Uppsala, Sweden), originally constructed for evaluation of multiple allergy reactivities.31 32 A full description of the microarray technology was published previously.22 Sixteen citrullinated ACPA peptides were investigated (see online supplementary table 1). Reactivity against non-citrullinated arginine-containing control peptides were investigated in parallel for all peptides, except for the citrullinated collagen type II peptide CII359-369 (citC1), where the control peptide is an autoantigen in itself and where the conformational epitopes are destroyed by citrullination.18 Fluorescense intensities were normalised and expressed as arbitrary units.
Supplementary file 1
Statistics
As ACPA responses are non-normally distributed, non-parametric statistics were used throughout the study. Diagnostic sensitivity was calculated in two ways. In the first (henceforth referred to as gross reactivity), gross data were evaluated and the 98th percentile among the controls were used as cut-off. In the second, net ACPA reactivity defined as the difference in arbitrary units between citrullinated and arginine peptides were calculated, and the 98th percentile for that difference among the controls used as cut-off. Additive sensitivity and specificity was calculated either from the full cohort, or starting from the anti-CCP2 sensitivity of 64.46% (1821/2825 patients positive) and specificity of 98.37% (9/551 controls positive) and thereafter adding the ACPA peptide that added most patients. Thereafter, the process was reiterated seven times, each time defining the ACPA specificity with the highest prevalence in the remaining group, and finally performed for all 16 peptides. For the calculation of the additive sensitivity and specificity for multiple ACPA reactivities as well as for determination of SE and smoking association among anti-CCP2 negative subjects, an alternative 99.5% specificity cut-off was evaluated in parallel. SE and smoking associations were expressed as OR with 95% CIs. Statistics were performed using the JMP11 and Prism 6 softwares.
Results
Use of arginine-subtracted ACPA data conveys higher diagnostic sensitivity and stronger association to SE alleles
To address the question concerning whether correction for background reactivity against arginine peptides would enhance the performance of the assay, the diagnostic sensitivity for individual ACPA peptides was evaluated both using gross and arginine-subtracted data. Out of the 15 peptides investigated in both ways, 12 showed higher diagnostic sensitivity when using arginine-subtracted data. There was a sizeable difference in sensitivity gain by using arginine-subtracted data, with total sensitivity for the two peptides Pept Z2 and Fibα621–635 increasing 10.55% and 7.36%, respectively. The mean increase in diagnostic sensitivity for all ACPA specificities was 3.20% (table 1). For the three peptides where raw data showed the highest sensitivity, the difference was negligible (0.11%–0.25%). Co-occurrence of reactivity against individual citrullinated peptides is described in online supplementary table 2.
Comparison of diagnostic sensitivity for 16 ACPA peptides between raw data for citrullinated peptides and net data where the responses for the corresponding arginine-containing control peptides had been subtracted from patients and controls before calculation
We then compared the individual association to SE for 15 peptides using both gross and arginine-subtracted data among the 2825 patients with RA, for each comparison using the negative patients as controls. The arginine-subtracted data yielded higher OR for SE for 12/15 peptides, whereas only 3/15 showed higher OR when gross data were used (table 2). The three peptides showing marginally decreased sensitivity after arginine subtraction (table 1) were also the same showing a marginal decrease in OR. The differences in OR were however small and overlapping between raw and arginine-subtracted data (table 2).
HLA-DRB1* shared epitope (SE) association for 16 citrullinated peptides among patients with RA from the EIRA cohort
We hypothesised that the increase in sensitivity after subtraction of arginine peptide reactivity might be due to differences between patients and controls concerning reactivity against arginine peptides. Indeed, as shown in (see online supplementary table 3), controls showed higher reactivity to 13/15 arginine-containing peptides than did patients with RA. Thus, individual subtraction of arginine peptide values increased the difference between mostly ACPA-positive patients and mostly ACPA-negative controls.
As arginine-subtracted data both increased the diagnostic sensitivity and association to HLA SE among patients with RA, arginine-subtracted data have been used below.
Increase in sensitivity and decrease in specificity by sequentially adding individual peptides to the assay
The individual diagnostic sensitivity for the evaluated peptides are shown in table 1 and varied between 16.53% and 57.73% after arginine subtraction. For the citrullinated type II collagen peptide CII359-369 where no arginine subtraction was performed, the sensitivity was 9.03%.
Thereafter, combinations of individual peptides were evaluated concerning their propensity to identify patients in the full RA group and within the anti-CCP2-negative RA subset. When using 98th percentile cut-offs, sequential addition of peptides increased the sensitivity from 57.73% for 1 peptide to 74.69% for 8 peptides and 77.95% for all 16 peptides. This increase in sensitivity was associated with a drop in diagnostic specificity from 98.19% for one peptide, 87.11% for eight peptides and 80.22% for all 16 peptides. When using 99.5% specificity cut-offs, addition of eight peptides increased the total sensitivity to 66.65% with maintenance of a rather high specificity (97.46%). For all 16 peptides, sensitivity was 68.04% and specificity was 95.10% (figure 1A, B).
Using the 98th percentile cut-off, sequential addition of peptides increased the anti-CCP2 sensitivity (64.46%) to 76.04% for eight added peptides and to 78.69% for all 16 peptides; the corresponding specificities were 98.37% (anti-CCP2 only), 86.75% and 79.67%. With 99.5% percentile cut-offs, addition of eight peptides increased the total sensitivity to 70.09% with 95.64% specificity. For all 16 peptides, sensitivity was 70.73% and specificity was 94.01% (figure 1C, D).

Impact on total (A and C) diagnostic sensitivity and (B and D) diagnostic specificity by sequentially adding the peptide adding most patients. In (A) and (B), all patients were investigated, and in (C) and (D), the anti-CCP2 negative subset. Sensitivity (64.46%) and specificity (98.37%) for anti-CCP2 are marked as horizontal dotted lines. In each panel, two optional lines are shown, based on 98% and 99.5% diagnostic specificity for the individual peptides, determined from 551 healthy controls. In panels (A) and (B), the order of the first eight peptides are Fibβ60-74, Pept-5, Fibβ36-52, Eno5-21, Fibα36-50, Fibα621-635, Pept-1 and Fil307-324 at the 98% specificity level, and Pept-5, Fibβ60-74, Pept-1, Pept-Z1, Fibβ36-52, Eno5-21, Fibβ621-635 and Fil307-324 at the 99.5% specificity level. In panels (C) and (D), the corresponding orders were Fibβ60-74, Fibα36-50, Pept-5, Fibα621-635, Fibβ36-52, Fibα580-600, Pept-1 and Pept-Z1 at the 98% specificity level, and Fibβ60-74, Pept-5, Pept-1, Fibβ36-52, Fibα36-50, Fibα621-635, Eno5-21 and Bla-26 at the 99.5% specificity level. CCP2, cyclic citrullinated peptide 2.
The distribution of number of ACPA specificities is shown in online supplementary figure 1. All patients analysed together showed a bimodal distribution with maxima around 0 and 13 specificities respectively (mean 6.27). This distribution was explained as an overlap of anti-CCP2 positive with mode 13 and mean 9.21 specificities and anti-CCP2-negative patients with mode 0 and mean 0.94 specificities. The low mean for the anti-CCP2-negative patients was however more than three times higher than for the healthy controls with mean 0.30 specificities. Only two controls had more than 4 ACPA specificities, 11 and 14 respectively, and both were anti-CCP2 positive.
Individual peptide reactivity and association to SE and smoking
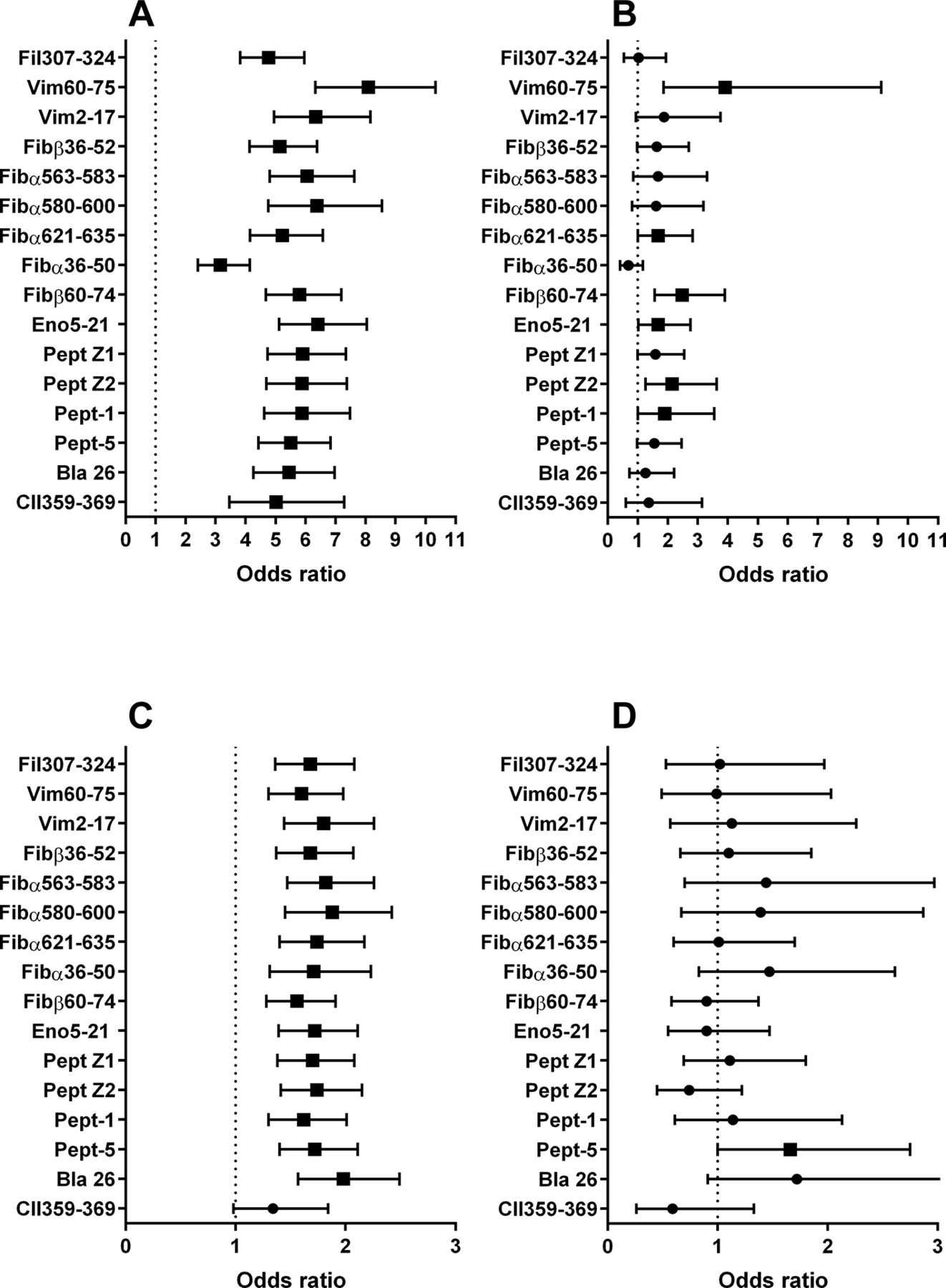
When RA patients showing reactivity to individual ACPA peptides were compared with patients not reacting with any ACPA peptides, all peptides showed a significant association to SE and all but one associated to a history of ever smoking. The OR for SE association varied between 3.16 and 8.09 (mean OR 5.69, 18.0% coefficiency of variation (CV%)) and was substantially higher than for the association to smoking (OR between 1.34 and 1.98, mean OR 1.71, 8.4 CV%). We thereafter investigated the SE and smoking associations among the anti-CCP2-negative patients for individual peptides. Six out of 16 peptides showed an individual SE association (mean OR for all 16 peptides 1.76, 40.4 CV%), and only 1/16 associated with smoking (mean OR 1.14, 27.7 CV%) and OR was generally much lower and showed higher variability than for the full RA cohort (figure 2 and online supplementary table 4).
The number of ACPA specificities varied with occurrence of SE, with a mean of 3.45 ACPA peptides for SE-negative and 7.28 for SE-positive patients (see online supplementary figure 2). Number of ACPA also related to the number of SE alleles; SE heterozygous patients showed a bimodal distribution very much like the total RA population with a mean of 6.59 peptides, while SE homozygous individuals showed a clear mode around 12, with a mean of 8.67 reactivities. When two-way analysis of variance (ANOVA) was applied using anti-CCP2 and SE status as independent variables, both associated to number of ACPA peptides (p<0.0001 for both), with a significant interaction (p=0.0010, data not shown).
A history of smoking also associated with higher number of ACPA specificities: never smokers reacted with a mean of 5.14 peptides, whereas ever smokers had 6.79 peptide reactivities as a mean (see online supplementary figure 2). Although this smoking association partly depended on the co-occurrence with HLA SE, a significant difference (p=0.0035) remained between the number of ACPA specificities among SE negative never smokers (mean 2.59, median 1) and SE negative ever smokers (mean 3.90, median 1; data not shown). Two-way ANOVA confirmed that number of ACPA specificities associated with both ever smoking status and SE (p<0.0001 for both), but without any interaction (p=0.55, data not shown).
Among anti-CCP2-negative patients HLA SE and smoking act in synergy as risk factors for ACPA peptide-positive but not for ACPA peptide-negative RA
We next investigated whether ACPA peptide reactivity associate with RA risk independent of anti-CCP2. Anti-CCP2-negative subjects (1004 patients with RA and 542 controls) were investigated in relation to SE and smoking history. As the combination of any of 16 peptide reactivities at the 98% specificity level creates a very low specificity (figure 1), we defined ≥2 peptide reactivities as the cut-off for general ACPA peptide positivity, in agreement with a previous publication.33 Occurrence of SE together with smoking history associated with significantly increased OR for RA (OR 3.55, CI 1.24 to 10.14), but only among ACPA-positive subjects (figure 3A, B and table 3). Also, using the 99.5% cut-off for individual peptides, SE and smoking history in synergy associated with RA risk among ACPA peptide positive (OR 3.86, CI 1.08 to 13.83) but not among ACPA-negative subjects in the anti-CCP2-negative subgroup (figure 3C, D and online supplementary table 5).

Individual association between (A and B) HLA-DRB1* shared epitope or (C and D) smoking with the occurrence of antibodies against 16 citrullinated peptides among 2825 RA patients. Analysis was performed for both (A and C) all patients and (B and D) the anti-CCP2-negative patient subset (n=1004). Arginine control values have been subtracted before calculation for the first 15 peptides, and raw data have been used for the collagen type II peptide CII359-369. Cut-off was set at the 98th percentile for 551 healthy controls. Results are shown as ORs with 95% CIs. ORs for significant associations are depicted as filled quadrants and for non-significant associations as filled circles. The corresponding figures are shown in online supplementary table 4. CCP2, cyclic citrullinated peptide 2.

{kind=link}
{kind=link}
{kind=link}
ORs for developing RA among anti-CCP2 negative subjects in relation to HLA-DRB1* SE alleles and a history of smoking, including 1004 RA patients and 542 healthy controls. In (A) and (B), ACPA positivity was defined as reacting with ≥2 peptides where the cut-off for each individual peptide was set at the 98th percentile of 551 healthy controls, and in (C) and (D) ACPA was defined as reacting with ≥1 peptide at the 99.5% specificity level. In (A) and (C) data are shown for ACPA peptide positive patients and in (B) and (D) data for ACPA peptide negative individuals are depicted. Significant OR with CI not involving one are shown with an asterisk. Full statistical data are shown in table 3 for panels (A) and (B), and online supplementary table 5 for panels (C) and (D). ACPA, anticitrullinated protein/peptide antibodies; CCP2, cyclic citrullinated peptide 2; RA, rheumatoid arthritis; SE, shared epitope.
Association to RA diagnosis for reactivity against 16 citrullinated peptides, HLA SE and smoking among anti-CCP2-negative subjects
When using a 98% specificity cut-off for individual peptides and regarding ≥1 reactivity as positive, the corresponding association remained barely significant (OR 1.86; CI 1.02 to 3.39; see online supplementary table 6).
Discussion
In this study we have shown that our ACPA microarray defines an ACPA-positive subgroup among anti-CCP2-negative patients that shows the same HLA SE/smoking association as previously described for anti-CCP2-positive patients. Indeed, our figure 3 on anti-CCP2-negative subjects dichotomised according to ACPA reactivity mimics a previously published figure on unselected patients with RA from EIRA dichotomised according to anti-CCP2 status although with smaller statistical effect.3 Therefore, it is plausible that the use of additional peptides can diagnose an extended group of ACPA-positive patients that will respond to therapies like anti-CCP2-positive patients.
Individual sample correction for reactivity against arginine peptides both increases diagnostic sensitivity and slightly increases association to SE among the patients. As shown in online supplementary table 3, arginine peptide reactivities were higher among controls than among patients with RA for most peptides, thus explaining a larger difference between mostly ACPA-negative controls and mostly ACPA-positive patients after individual subtraction of arginine peptide reactivity. In our previous proof of concept study, we raised the question on whether subtraction of reactivity against arginine peptides would enhance assay performance.22 Including more peptides and patients, we, this time, obtained quite unequivocal results showing that arginine subtraction yields a considerable increase in diagnostic sensitivity at an unchanged specificity level, together with increased association to SE among patients with RA for the majority of peptides. The microarray format easily allows such sample specific controls to be performed. Among routine clinical diagnostic tests, very few ACPA assays include such sample-specific normalisation, as that would imply almost the double number of ELISA wells needed and thus a considerable increase in cost per patient analysis.
Sample-specific normalisation is however not generally used in other ACPA multiplexing formats, for example, in the recently published study by Wagner et al 33 using addressable laser bead immunoassay.
In a recent study by van Heemst et al 34 using the same assay but considerably fewer samples, no association was found between SE and smoking among anti-CCP2 negative subjects. In that study, the mean +2 SDs among controls were used as cut-off for each peptide in an additive manner, thus creating a considerably lower total specificity. When using a comparable low-specificity approach, also our corresponding results became considerably weaker, although our study includes >5 times more patients with SE data than the Leiden study. We conclude that to be meaningful, microarrayed ACPA tests have to be combined with high specificity approaches.
In our study, 118 samples were excluded due to specific properties of the individual sera, including general sticky binding to the microarray surface and general background reactivity to all streptavidin-containing spots. In the absence of individual controls and control over the full reaction surface, such sera would probably be regarded as positive for the measured analyte. We found only two controls with a high number of ACPA reactivities, and both belonged to the minority (9/551) of anti-CCP2-positive controls. Hypothetically, these two individuals might be in a pre-RA phase, as the number of ACPA peptide reactivities increase during the years before RA diagnosis.35 We believe that our assay format helped us to exclude non-assessable samples, with ensuing clear difference in frequency distribution between anti-CCP2-positive and anti-CCP2-negative patients as shown in online supplementary figure 1B and C.
Using the eight most discriminatory peptides and high individual specificity (99.5%), the total sensitivity was increased by 5.5% as compared with anti-CCP2 alone, with the preservation of total specificity of 95.64%. This implies that 16% of the anti-CCP2-negative patients can be diagnosed as ACPA positive with a total specificity in agreement recommendations for rheumatoid factor in the previous ACR classification criteria (>95%), which is the only directive so far for cut-off setting for RA autoantibodies.29
This assay has been used to study the sequential appearance of individual ACPA reactivities the years predating RA.35 We envisage that this assay can be used to study which ACPA reactivities might appear in patients with arthralgia and in first-degree relatives of patients with RA as has previously been done with isotypes of anti-CCP2 and rheumatoid factor.36
In conclusion, our study demonstrates that the multiplex assay can identify a substantial proportion of anti-CCP2-negative patients that display ACPA reactivities, and in addition that this subset of patients with RA shows similarities with the classical anti-CCP2-positive RA group concerning major genetic and environmental determinants. This extended group of ACPA-positive patients will most likely use similar molecular pathways towards disease and respond similar to therapies as anti-CCP2-positive patients where therapeutic responses are contingent on ACPA status.
Acknowledgments
The authors would want to thank the EIRA study group for including patients. This study was supported by grants from the Swedish Research Council, the Swedish Rheumatism Association, King Gustav V 80-year Foundation and the Be The Cure EU consortium.
References
Footnotes
Handling editor Tore K Kvien
Contributors JR had full access to all data, is responsible for data integrity, made statistical calculations and drafted the manuscript. JR, MH, LM-A and LK conceived the study. LA provided epidemiological data and statistical expertise. MH and LM-A participated in the development of the ISAC microarray and performed the laboratory work on the EIRA cohort. MC, ER, P-JJ, RH, KS, GS and KL provided peptides for the analyses, including validation of their performance. All authors read, commented on and approved the final manuscript.
Competing interests The project is part of the Innovative Medicines Initiative (IMI) project Be The Cure where Karolinska Institutet is a scientific partner and Thermo Fisher Scientific is a commercial partner. The project follows the rules for IMI projects. JR has obtained reagents from Thermo Fisher Scientific for the investigation on other rheumatology cohorts. LM-A is employed by Thermo Fisher Scientific. LK and RH were cofounders of a company, Curara AB, which has previously collaborated with Thermo Fisher Scientific concerning certain technical aspects of the multiplex assay. This development is done with partial support from an ERC Proof of Concept Grant (pRActice) to LK. RH is a coinventor of a patent (US Patent 7148020) protecting the use of the CitC1 and C1 peptides. KS is coinventor of the patents US 13/141,960 and EP 09799354.7 describing the diagnostic use of the hnRNP-A3 peptide epitopes. GS is coinventor of several international patents about ACPA antigens held by BioMérieux Cy and licenced to Eurodiagnostica Cy and Axis-Shield Cy for commercialisation of the CCP2 assays; according to French laws, he receives a part of the royalties paid to the Toulouse III University and the University Hospital of Toulouse. KL is coinventor of patent US12/524,465, describing the diagnostic use of the CEP-1 epitope. The other authors declare that they have no competing interests.
Ethics approval The ethics board in Stockholm.
Provenance and peer review Not commissioned; externally peer reviewed.
Data sharing statement All relevant data are available in the manuscript and supplementary files.