Article Text

Abstract
Objectives Familial aggregation of primary Sjögren’s syndrome (pSS), systemic lupus erythematosus (SLE), rheumatoid arthritis (RA) and co-aggregation of these autoimmune diseases (ADs) (also called familial autoimmunity) is well recognised. However, the genetic predisposition variants that explain this clustering remains poorly defined.
Methods We used whole-exome sequencing on 31 families (9 pSS, 11 SLE, 6 RA and 5 mixed autoimmunity), followed by heterozygous filtering and cosegregation analysis of a family-focused approach to document rare variants predicted to be pathogenic by in silico analysis. Potential importance in immune-related processes, gene ontology, pathway enrichment and overlap analyses were performed to prioritise gene sets.
Results A range from 1 to 50 rare possible pathogenic variants, including 39 variants in immune-related genes across SLE, RA and pSS families, were identified. Among this gene set, regulation of T cell activation (p=4.06×10−7) and T cell receptor (TCR) signalling pathway (p=1.73×10−6) were particularly concentrated, including PTPRC (CD45), LCK, LAT–SLP76 complex genes (THEMIS, LAT, ITK, TEC, TESPA1, PLCL1), DGKD, PRKD1, PAK2 and NFAT5, shared across 14 SLE, RA and pSS families. TCR-interactive genes P2RX7, LAG3, PTPN3 and LAX1 were also detected. Overlap analysis demonstrated that the antiviral immunity gene DUS2 variant cosegregated with SLE, RA and pSS phenotypes in an extended family, that variants in the TCR-pathway genes CD45, LCK and PRKD1 occurred independently in three mixed autoimmunity families, and that variants in CD36 and VWA8 occurred in both RA-pSS and SLE-pSS families.
Conclusions Our preliminary results define common genetic characteristics linked to familial pSS, SLE and RA and highlight rare genetic variations in TCR signalling pathway genes which might provide innovative molecular targets for therapeutic interventions for those three ADs.
- autoimmune diseases
- T cells
- rheumatoid arthritis
- Sjøgren's syndrome
- systemic lupus erythematosus
Statistics from Altmetric.com
Key messages
What is already known about this subject?
It is well recognised that rheumatoid arthritis (RA), primary Sjögren’s syndrome (pSS) and systemic lupus erythematosus (SLE) cluster in families, indicating the presence of shared variants underlying the genetic predisposition to familial RA, pSS and SLE. Genome-wide association studies have identified common variants (minor allelic frequency >1%) shared among RA, pSS and SLE.
What does this study add?
This study comprehensively characterised the genetic profiles across RA, pSS and SLE families, and further investigated the genetic background for their familial clustering. Our data identified certain rare and potential pathogenic variants concentrated in the T cell receptor (TCR) signalling pathway.
How might this impact on clinical practice or future developments?
The TCR signalling pathway genes found in this study may provide novel molecular targets for therapeutic interventions for RA, pSS and SLE, which remain a challenge to manage clinically.
Introduction
Rheumatoid arthritis (RA), systemic lupus erythematosus (SLE) and primary Sjögren’s syndrome (pSS) are three closely related autoimmune diseases (ADs), which share multiple disease aspects, including epidemiological characteristics, clinical manifestations and serological profiles. Indeed, SLE, RA and pSS co-occurring at the same or at different times in the same patient support the concept that these disorders share common molecular ties.1 Furthermore, co-aggregation of RA, SLE and pSS within family, also known as familial autoimmunity, indicates that common germline variants may predispose to these ADs.2 Previously, several associated genes shared among RA, SLE and pSS were identified by whole genome-wide association studies, including HLA, IRF5, STAT4 and TNFAIP3.3 4 Recently, whole-exome sequencing also has been used to uncover rare variants for SLE and RA.5 6 Nevertheless, rare variants for familial AD still remain an unsettled question.
Personalised therapies based on individual genetic susceptibility may increase the probability of improved efficacy.7 Harnessing the plasticity of CD4+ T cells could provide improved therapeutic efficacy for patients with AD.8 CD4+ T cells, including Th1, Th2 and Th17 cells, were identified in target organ infiltrates and peripheral blood in early ADs. It was also shown that CD4+ T cells play a major role in AD pathogenesis, amplifying inflammation by secretion of pro-inflammatory cytokines plus helping B cells to generate autoantibodies.9 However, the contribution of genetic factors to inappropriate T cell homing of Th1, Th2 and Th17 cells in AD still unknown.
Herein, we describe rare and deleterious variants in genes involved in TCR signalling pathway genes underlying predisposition to familial ADs.
Patients and methods
Family ascertainment
Subjects met research classification criteria using the 2002 American-European Consensus Group for pSS, the 2012 Systemic Lupus Collaborating Clinics for SLE and the 2010 ACR/EULAR for RA. Familial RA, pSS, SLE and autoimmunity were defined as patients with at least one first-degree relative with confirmed RA, pSS or SLE. Voluntary, written, fully informed consent was obtained from all participants.
For more details about experimental and bioinformatic methods, see the online supplementary methods and figure S1.
Supplemental material
Supplemental material
Supplemental material
Results
Genetic spectrum and gene enrichment for T cell-mediated immunity
Our filtering pipeline showed that pathogenic variants cosegregated with phenotypes on a per-family basis. We did not identify any significant linkage peak and clustering of variants located in genomic ‘hotspots’. Depending on family size, between 1 and 50 variants were shared by all affected individuals within each family (see online supplementary figure S2 and table S2), including 39 immune-related genes (table 1 and figure 1A). Previously implicated autoimmune-associated genes with new rare variants were identified, such as CFHR4, RPP21, IL12A and ARL14.10 To further dissect the shared genetic spectrum across families, we assessed whether there were enriched gene ontology terms within our immune-related gene set. Notably, regulation of T cell activation was highlighted (p=4.06×10−7, figure 1B, D), indicating that T cells probably contribute to the genetic basis of AD via different genes and/or pathways.
Supplemental material
Supplemental material
List of candidate genes prioritised by immune-related function and enrichment in T cell receptor signalling pathway

Results of whole-exome sequencing revealed T cell-mediated autoimmune. (A) Functional category of immune-related gene set identified by whole-exome sequencing. (B) Gene ontology analysis suggested T cell activation was a functional category by REVIGO scatterplot. (C) KEGG pathway enrichment indicated T cell receptor (TCR) pathway was significantly highlighted. (D) Metascape network of the enriched gene ontology terms showed the T cell activation was the central node. (E) Genetic variants in TCR pathway genes. Genes with variants were highlighted in yellow colour and italic type.
Sharing TCR-pathway genes across RA, SLE and PSS families
The immune-related genes were further classified according to their potential roles in signalling pathways. T cell receptor (TCR) signalling pathways (p=1.73×10−6) were aggregated (figure 1C), including PTPRC (CD45), ITK, TEC, LAT, LCK, PAK2 and NFAT5. To identify more TCR-pathway genes which were not included in the current KEGG database, we manually filtered immune-related gene sets further and found additional TCR-proximal genes for THEMIS, PLCL1, PRKD1, DGKD and TESPA1 (figure 1E). Notably, six TCR-pathway genes (ITK, LAT, THEMIS, PLCL1, TEC and TESPA1) were found within the LAT–SLP76 complex.11 The positions and conservation in different species of these variants and their corresponding gene expression were shown. Furthermore, the T cell proliferation and cytokine production were significantly increased with mutant TCR-pathway genes (see online supplementary figures S3–S6).
Supplemental material
Supplemental material
Supplemental material
Supplemental material
TCR interactive genes across RA, SLE and PSS families
We also identified TCR-interactive genes which are cross-linked to the TCR-pathway, including P2RX7, CERK, LAG3, SEMA4D, IRAK4, LAX1, CD36 and DPP4 (CD26). LAG3 and PTPN3 inhibit the activation of the TCR pathway12 13 and SEMA4D–CD72 interactions were necessary for TCR-induced T cell proliferation; in contrast, CD36, CD26 and IRAK4 influenced TCR signalling via lipid rafts.14 15 Also, P2RX7 signalling affected TCR signalling at the αβ/γδ lineage bifurcation checkpoint.16 The detailed connection of these genes and the TCR-pathway are shown in table 1.
Overlapping genes across RA, SLE and PSS families
Almost all of the identified variants are present only once in our families, indicating private genetic profiles and polygenic characteristics. Five immune-related genes with fully segregating variants in more than one family revealed a common pattern of genetic variants across pSS, RA and SLE (figure 1C and table 1), including CD36, PTPRC, RIPK1, VWA8 and TRIM16.
CD36 was embedded in RA and pSS families and VWA8 was shared in two extended pSS families and SLE family; while variants in TCR-pathway gene CD45 occurred in pSS, RA and mixed autoimmunity families. Recurrent stop-gain variants (p.S48X) in TRIM16 were replicated in pSS families and reflect the same TRIM family as TRIM21 (Ro52/SSA), which had been demonstrated to promote antiviral immunity.
We also detected a DUS2 variant (p.R358C) which cosegregated with SLE, RA and pSS phenotypes (figure 2) in an extended family; DUS2 and interferon-induced protein kinase are involved in the regulation of innate antiviral immunity.17 This provided direct evidence for sharing genes in these disorders. The presence of the TRIM16 and DUS2 variants supported the hypothesis that antiviral immunity may contribute to the development of familial AD.18



{kind=link}
{kind=link}
{kind=link}
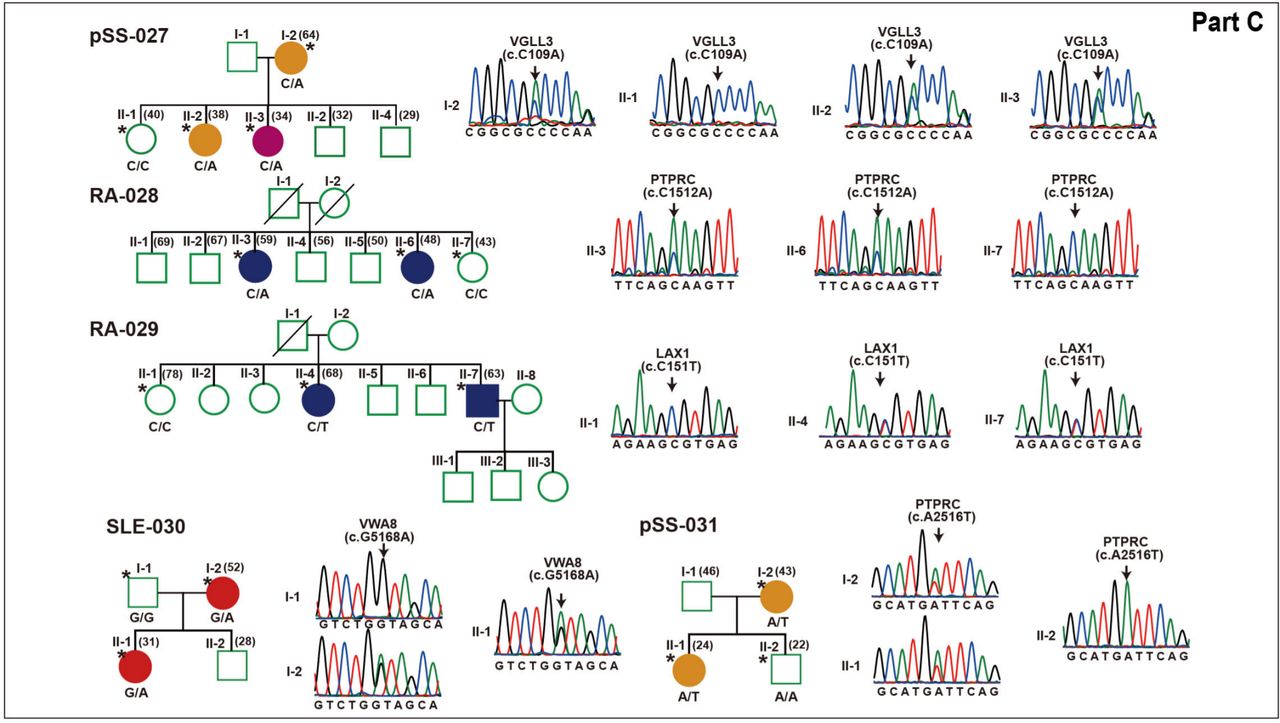{kind=link}
Rare variants segregating with phenotypes within each family. Pedigree structures of 31 families with genetic variants listed below each family member are shown. Males are indicated with squares, females with circles, slashes indicate deceased members and clinical diagnoses are shown by different colour. All subjects with DNA available are indicated by genotype, individuals with whole-exome sequencing are indicated by single asterisks and extended subjects for validation are indicated only by genotype. Sanger sequencing chromatograms are shown near the pedigrees.
Discussion
It is well known that RA, SLE and pSS cluster in families. However, not all of the genetic background that explains this clustering has been discovered and it remains unclear whether specific genetic variants and pathways are shared in familial SLE, RA and pSS. Our study describes genetic profiles across SLE, RA and pSS in Chinese families.
In fact, symptoms of SLE, RA and pSS at an early stage are similar, although tissue damage may intervene or features of one may dominate over another. We identified a single variant in the DUS2 gene responsible for part of SLE, RA and pSS phenotypes. CD45, LCK and PRKD1 in mixed autoimmunity families further supported the presence of some common genetic variants underlying aspects of SLE, RA and pSS.
Consistent with previous studies showing polygenic variants in SLE families,19 we identified 39 immune-related genes (the majority being T cell variants but not B variants), which supported the hypothesis that T cell-initiated immunity drives AD parthenogenesis. Our results show that genetic variants in T cells, but not in B cells, supply a genetic basis for AD in some familial forms of pSS, SLE and RA. We speculate that common variants of T cell activation differentiate and acquire distinct functions in subtypes which contribute to AD phenotypes and serological profiles via B cells.
TCR-pathway genes accounted for up to 48.7% of the immune-related genes in our AD families, suggesting SLE, RA and pSS could be influenced by multiple rare variants acting on a common pathway. A dynamic role for Th-cell activation is characterised by cytokine secretion, including Th1, Th2 and Th17, critical contributors to pSS, SLE and RA pathogenesis.20 21 In this study, variants we found in CD45, LCK, LAT, TEC and NFAT5 activate the coagulation cascades and the TCR signalling pathway and they might explain the triggering of naive CD4+ T cell differentiation in pSS, RA and SLE. Most of them were TCR proximal rheostats serving the LAT–SLP76 complex. We propose primarily variants linking genetic susceptibility and naive CD4+ T cell differentiation, amplified small initial difference in the efficacy of triggering in TCR signalling results the profound effects underlying AD pathogenesis. Furthermore, diverse AD presented in mixed autoimmunity families, strongly indicated genetic variants in TCR-pathway genes contributed to phenotypic heterogeneity.
Our results present variants in the TCR interactive genes leading to T cell activation, resulting in T cell differentiation, survival and effectors functions also contributed AD phenotypes. TCR-pathway and its interactive genes may not represent the full spectrum of AD susceptibility. The different genetic variants were underling not only AD susceptibility but also with specific AD phenotypes.
A total of five shared gene patterns added further data to help understand the root and specific genetic aetiologies of ADs. CD36 is a known susceptibility gene for malaria,22 and regulates malaria-induced pro-inflammatory cytokine responses and activation of T cells.23 P2XR7 drives the fine-tuning between Th1 and Tfh cell differentiation to protect against malaria.24 The antimalarial agent chloroquine has an established role and is routinely used in the treatment of RA, SLE and pSS. However, the common mechanism of action of this medication in malaria and ADs remains unclear. Therefore, we suspected that CD36 and P2XR7 might be novel immune links between malaria and ADs.
Currently, B cell depletion interrupts T cell/B cell interaction,25 and may lead to worsening of the AD.26 Characterisation of the TCR gene might also open new avenues to design specific and more effective therapies. Our study provides potential therapeutic targets, such as inhibitors AX-024 for the LAT–SLP76 complex,25 ITK inhibitor ibrutinib27 and TEC inhibitor PRN694.28 Overlapped immune-related genes also could be severed as therapeutic targets, such as GSK2982772 for RIP1K-I40 for CD26.29
In conclusion, the families presented here identified common genetic patterns across RA, SLE and pSS, and provide data on the genetic background for SLE, pSS and RA which may help understand their pathogeneses. They also illustrate an important concept that could explain some of the heterogeneity in SLE, pSS and RA families. Further studies are warranted to evaluate the consequences of TCR-pathway variant and the mechanism by which it contributes to the development of familial ADs.
Acknowledgments
The authors thank the patients and their family members for participation in the study.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Footnotes
Handling editor Josef S Smolen
YW and SC contributed equally.
Correction notice This article has been corrected since it published Online First. Figure 2 has been amended.
Contributors GZ and YW designed the study. YW, JC, XX, SG, CZ, JW, JL, QL and SZ performed experiments and analysed data. GZ, DEF and MM-C wrote the manuscript. SC and RM helped with sample collection.
Funding Supported by grants from the Natural Science Foundation of Guangdong Province (2014A030307003).
Competing interests None declared.
Patient consent for publication Not required.
Ethics approval The Ethics Committee of Shantou Central Hospital.
Provenance and peer review Not commissioned; externally peer reviewed.
Data availability statement Data are available in a public, open access repository.