Article Text

Abstract
Objectives: Monitoring of peripheral B-cell subsets in patients with systemic lupus erythematosus (SLE) revealed an activity-related expansion of CD27++CD20−CD19dim Ig-secreting cells. A similar subset has also been identified 6–8 days after tetanus/diphtheria vaccination in normal individuals and in patients with infectious disease.
Methods: This subset was analysed further focussing on the HLA-DR surface expression in a cohort of 25 patients with SLE.
Results: This study revealed that 86% (range 59–97%) of CD27++CD20−CD19dim cells express high levels of HLA-DR, are also expanded in the bone marrow, and represent plasmablasts enriched with anti-dsDNA secreting cells. The remaining CD27++CD20−CD19dim cells were HLA-DRlow and represent mature plasma cells. Importantly, HLA-DRhigh plasmablasts showed a closer correlation with lupus activity and anti-dsDNA levels than the previously identified CD27++CD20−CD19dim cells.
Conclusion: HLA-DRhighCD27++CD20−CD19dim plasmablasts represent a more precise indicator of lupus activity and suggest that there is an overproduction or lack of negative selection of these cells in SLE.
Statistics from Altmetric.com
In addition to defective tolerance mechanisms accounting for the survival of self-reactive B cells in systemic lupus erythematosus (SLE) an increased activation of B cells has repeatedly been reported. In this context, co-stimulatory molecules have been found to be overexpressed on lupus T and B cells.1 2 3 Whereas the exact mechanisms involved in B-cell activation, including T-cell-dependent and independent pathways leading eventually to a secretion of characteristic autoantibodies in lupus are highly complex and remain largely unknown, this study focussed on the characterisation of the immediate product of aberrant B-cell activation—a CD27++CD20−CD19dim Ig-secreting cell subset. These cells have previously been identified in the peripheral blood of patients with SLE, and were shown to correlate with disease activity.4 5 In normal individuals, only 1–2% of all peripheral blood CD19+ cells are CD27++CD20−CD19dim.5 However, this subset has also been demonstrated to expand temporarily 6–8 days after secondary immunisation with tetanus toxoid,6 comprising an antigen-specific fraction with high HLA-DR expression. These freshly generated antigen-specific plasmablasts migrated towards SDF-1 and were considered to be destined to home in the bone marrow. The transcription of HLA-DR has been shown to be downregulated during plasma cell development as a consequence of increased BLIMP-1-levels.7
So far, CD27++CD20−CD19dim B cells have only been characterised in few lupus patients with regard to HLA-DR expression5 focusing on their characterisation as Ig-producing cells. However, it appears to be of critical importance to analyse the ratio of HLA-DRhigh/HLA-DRlow cells in patients with SLE in order to differentiate recently generated plasmablasts from mature plasma cells. In order to address this question, we performed a cross-sectional analysis of 25 lupus patients. The results suggest that recently generated HLA-DRhigh plasmablasts predominate over HLA-DRlow plasma cells in the peripheral blood of lupus patients and that this bias is dependent on disease activity. HLA-DRhigh plasmablasts were also found in bone marrow aspirates obtained from patients with SLE consistent with their influx into the bone marrow. However, it remains to be elucidated whether and to what extent they may be able to mature into plasma cells, replace established plasma cells or undergo apoptosis in individual patients.
The data are consistent with an activity-related, increased generation of HLA-DRhigh plasmablasts in SLE.
Materials and methods
After informed consent was obtained, peripheral blood was drawn from 25 (24 female, 1 male) lupus patients hospitalised at our unit (Charité University Hospitals, Berlin, Germany). Patient characteristics are shown as supplementary material (available online only). In addition all methods are described in the supplement (supplementary text file, available online only).
Results
In the current lupus patient cohort, 12.2% (SD 10.6) of all circulating B cells were determined to be CD27++CD20−CD19dim. This subset has previously been found to be expanded in lupus patients comprising plasmablasts and plasma cells.4 5 As illustrated in fig 1, this expansion was mainly related to an increase of HLA-DRhigh plasmablasts (median 86%; range 59–97%) compared with HLA-DRlow plasma cells. Interestingly and similar to peripheral blood, HLA-DRhigh plasmablasts were also found to be expanded in the bone marrow of two lupus patients (fig 1). In contrast, only a low percentage of HLA-DRhigh plasmablasts was found in bone marrow from healthy controls (1–1.5% of CD19+ cells or <0.1% of bone marrow mononuclear cells, respectively, fig 1).

(A) Gating strategy. Peripheral blood mononuclear cells were labelled with fluochrome-tagged antibodies to CD19, CD20, CD27 and HLA-DR. After gating on CD19+ cells, CD27++CD20− cells were gated (plot) and HLA-DRhigh and –low subsets (histogram) were analysed. The blue line shows the expression of HLA-DR, gated on the CD27++CD20− subset isolated from the peripheral blood, for comparison the corresponding bone marrow (BM) subset is shown in grey. (B) Expression of HLA-DR on peripheral blood (PB) and bone marrow B cells and plasmablasts as well as plasma cells in a representative patient with systemic lupus erythematosis (SLE). For comparison, the result of peripheral blood from a normal donor (ND) and the bone marrow from another normal subject is also shown.
Further analyses sought to investigate if any of these circulating CD27++CD20−CD19dim subsets correlate with disease activity in lupus patients. As a result, we identified a significant correlation between the absolute number or frequency of HLA-DRhigh plasmablasts and the disease activity as assessed by the systemic lupus erythematosus disease activity index (SLEDAI) score (p<0.04 and p<0.004, respectively) or IgG anti-double-stranded DNA antibody levels (p<0.03, p<0.01, respectively) in 25 patients with SLE (table 1). Interestingly, analysis of the entire CD27++CD20−CD19dim subset revealed a correlation between the absolute cell number and lupus activity (p<0.005) and between its frequency and anti-dsDNA titres (p<0.03) but no other significant correlations. In contrast, HLA-DRlow plasma cells, total CD27+ memory B cells as well as CD27− B cells did not correlate with disease activity or IgG anti-dsDNA antibody levels. These data clearly support that HLA-DRhigh plasmablasts are the immediate product of immune activation in lupus and that this subset is associated with disease activity more closely than the entire CD27++CD20−CD19dim subset.
Statistical relation between the frequency and absolute number of HLA-DRhigh plasmablasts, HLA-DRlow plasma cells as well as CD27+ and CD27− B-cell subsets with lupus activity recorded by the SLEDAI and IgG anti-dsDNA levels
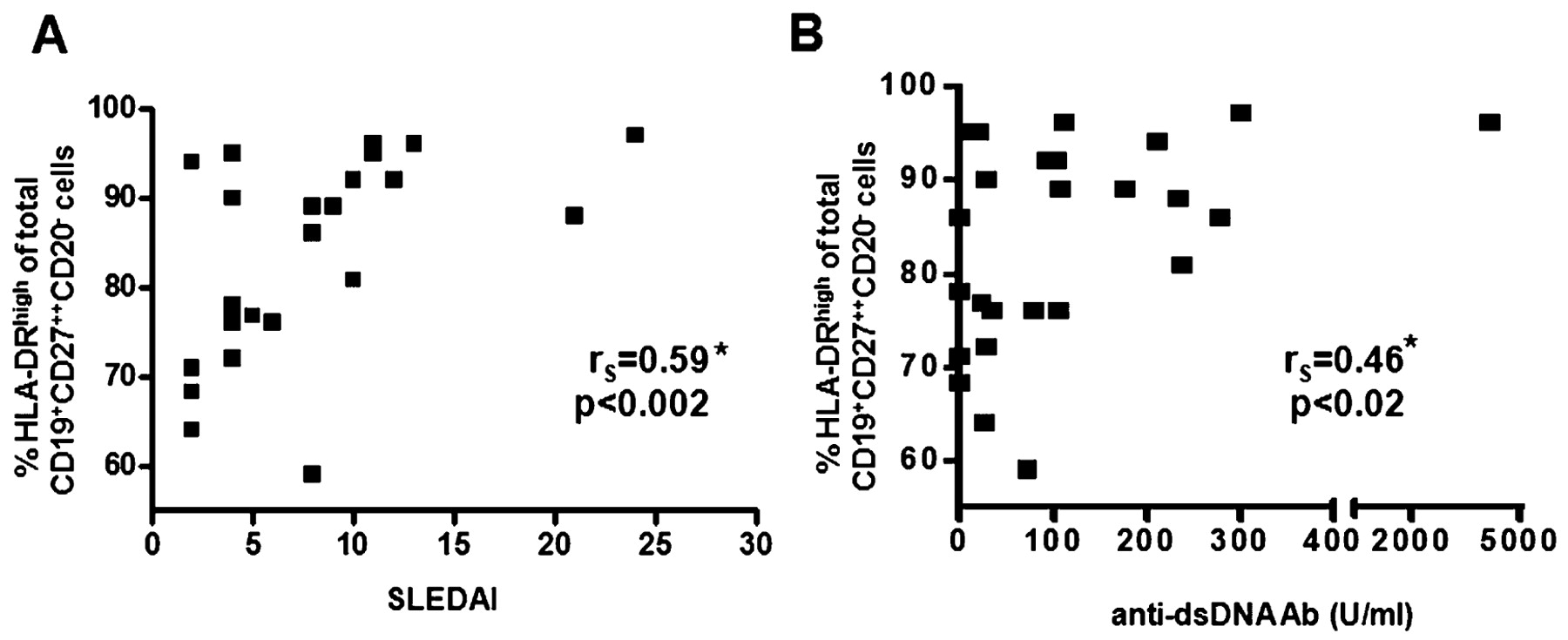
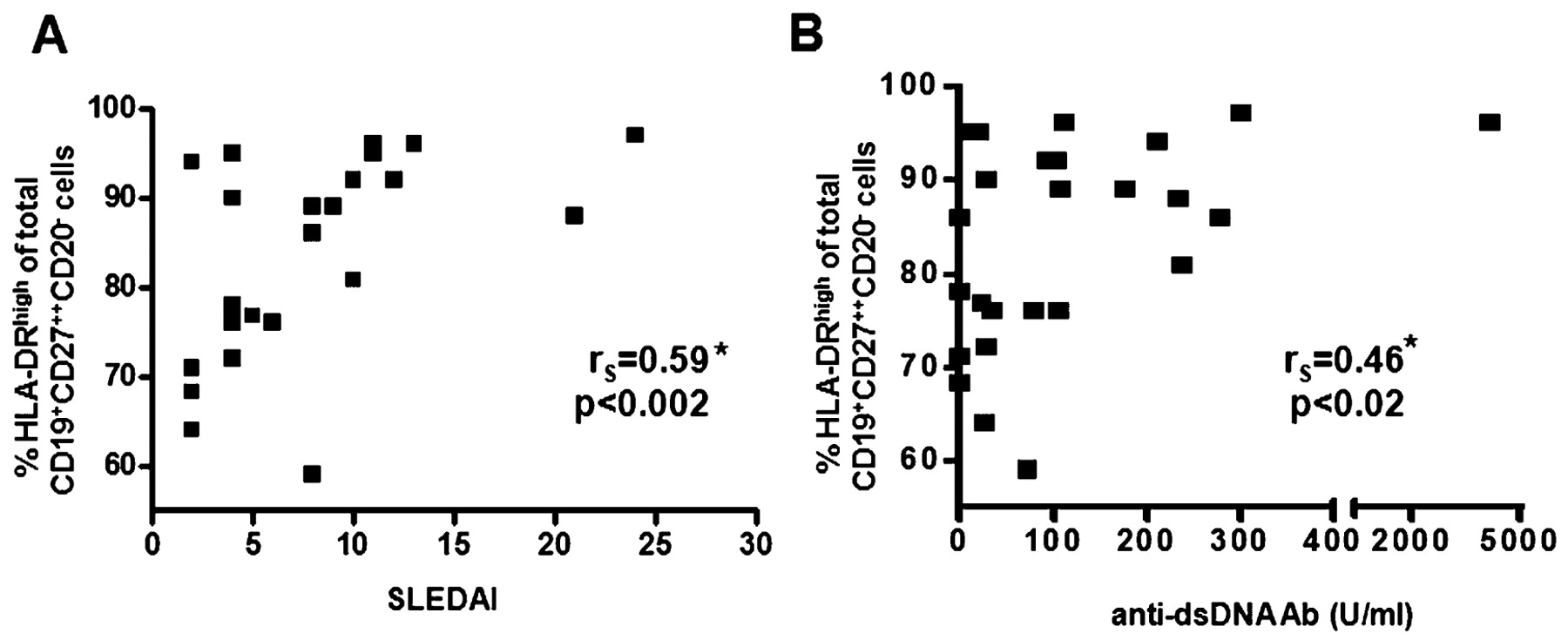
In line with these results, we found the percentage of HLA-DRhigh plasmablasts within the CD27++CD20−CD19dim subset to be significantly correlated with the SLEDAI score (rS = 0.59, p<0.002, fig 2A) and the level of IgG anti-dsDNA antibodies (rS = 0.46, p<0.02, fig 2B). To test whether HLA-DRhigh plasmablasts might be directly related to the IgG anti-dsDNA antibody serum levels, we analysed the frequency of IgG anti-dsDNA antibody-secreting cells among both the HLA-DRhigh plasmablast subset and HLA-DRlow plasma cell subset in a lupus patient. Of note, the frequency of IgG anti-dsDNA antibody-secreting cells was more than twice as high in the HLA-DRhigh plasmablast subset compared with the HLA-DRlow plasma cell subset (3.3% (5/153) vs 1.5% (3/198) of all IgG-secreting cells).
{kind=link}
{kind=link}
Significant correlation between the HLA-DRhigh subset of CD27++CD20−CD19dim cells (%) and disease activity assessed using the systemic lupus erythematosus disease activity index (SLEDAI) (A) and IgG anti-dsDNA antibody levels (B). *Spearman rank correlation.
Discussion
This study identified that among circulating CD27++CD20−CD19dim cells known to be enhanced in active patients with SLE,4 5 recently generated HLA-DRhigh plasmablasts predominate over HLA-DRlow plasma cells. Their absolute number and frequency correlate with disease activity more significantly than the entire CD27++CD20−CD19dim subset, which appears to be a mixed population that also contains HLA-DRlow plasma cells not related to disease activity. Although anti-dsDNA IgG antibody production could be identified in both subsets, there was an enhanced production by HLA-DRhigh plasmablasts derived from peripheral blood further supporting their pathogenic role and relation with disease activity. Freshly generated HLA-DRhigh tetanus-specific plasmablasts have been found to be expanded in the peripheral blood of healthy individuals 6–8 days after secondary immunisation.6 In comparison, most circulating CD27++CD20−CD19dim cells (86%; range 59–97%) express high levels of HLA-DR in patients with SLE. However, the expansion of HLA-DRhigh plasmablasts does not appear to be restricted to the peripheral blood, because we could also readily identify them in bone marrow aspirates in two lupus patients. However, the source and fate of these HLA-DRhigh plasmablasts as well as the mechanisms involved in their trafficking remain unknown.
In this context, a previous study identified an enlarged population of plasma cells in kidneys from patients with lupus nephritis adjacent to germinal centre-like structures and local Ig production in the affected organ,3 as has been shown in lupus nephritis8 and the spleen9 in animal models. These models offer the advantage of addressing various questions more thoroughly. Feeding NZB/W mice with bromodeoxyuridine, Hoyer et al.9 documented substantial populations of long-lived plasma cells and short-lived plasmablasts populating the spleen, with only the latter being sensitive to cyclophosphamide treatment. In line with these experimental data, the abundance of HLA-DRhigh plasmablasts in different organs including peripheral blood and bone marrow is notable in patients. Whether these cells further differentiate into HLA-DRlow plasma cells and undergo specific selection processes or just reflect the characteristic overflow observed in the tissues of lupus patients remains to be elucidated.
Although a decline in HLA-DRhigh plasmablasts noted almost immediately after methylprednisolone treatment as well as 2 months after B-cell depletion were noted in individual patients (supplementary fig 1, available online only), longitudinal studies of large cohorts of patients are required to assess the effect of particular drugs on HLA-DRhigh plasmablasts. Several factors contributing to B-cell activation and differentiation into plasmablasts or plasma cells have been identified in patients with SLE and other autoimmune diseases. In this context, T-cell and antigen-dependent and independent pathways, certain cytokines and B-cell activating factor have been implicated to drive plasma cell differentiation from memory, naive and even transitional B cells in humans.10 It is thus also possible that the expansion of HLA-DRhigh plasmablasts in lupus results from premature B-cell activation and differentiation. In this regard, CD27−IgD−CD95+ memory B cells showing an activated phenotype were found to be expanded in SLE patients with active disease.11
An important mechanism to control B-cell activation and differentiation at the post-germinal centre stage is mediated through FcγRIIb. Usually, FcγRII expression is upregulated on activated B cells and germinal centre cells.12 In contrast, memory B cells show a decreased surface expression of FcγRIIb associated with an impaired regulation of B-cell receptor signalling in a subset of patients with SLE,13 suggesting that the expansion of plasmablasts and autoantibody secretion in lupus patients might be related to an impairment of this post-germinal centre control mechanism, as also observed in autoimmune mouse models.14 15
In conclusion, expanded HLA-DRhigh plasmablasts represent a more precise marker of B-cell activation in SLE closely related to disease activity, including the production of IgG anti-dsDNA antibodies. The expansion of these cells reflects a lack of immune regulation and selection, respectively, which requires further studies.
REFERENCES
Supplementary materials
Web Only Data 69/1/305
Files in this Data Supplement:
Footnotes
▸ Additional supplementary material and supplementary fig 1 is published online only at http://ard.bmj.com/content/vol69/issue1
Funding Supported by grant Do 491/5-5 and in part by SFB 650/TP16, the Deutsches Rheumaforschungszentrum is supported by the Senate of Berlin Germany.
Competing interests None.
Ethics approval The local Institutional Review Board (Charité Berlin) approved the study.
Patient consent Obtained.