Article Text

Abstract
Objectives To study the presence of interferogenic autoantibodies in systemic sclerosis (SSc) and their correlation with clinical manifestations, serum levels of interferon α (IFNα) and chemokines of importance in the disease process.
Methods Peripheral blood mononuclear cells (PBMCs) or purified plasmacytoid dendritic cells (pDCs) from healthy donors were stimulated with sera from patients with SSc (n=70) or healthy individuals (n=30), together with necrotic or apoptotic cell material. The IFNα produced and serum levels of IFNα, IFN-inducible protein-10 (IP-10)/chemokine (C-X-C motif) ligand 10, monocyte chemoattractant protein-1 (MCP-1)/(C-C motif) ligand-2 (CCL-2), macrophage inflammatory protein-1α (MIP-1α)/CCL-3 and RANTES/CCL-5 were measured and correlated with the presence of autoantibodies and clinical manifestations in the patients with SSc.
Results Sera from both diffuse SSc and limited SSc contained interferogenic antibodies, which correlated with the presence of anti-ribonucleoprotein and anti-Sjögren syndrome antigen autoantibodies. The pDCs were responsible for the IFNα production which required interaction with FcγRII and endocytosis. Increased serum levels of IP-10 were associated with vascular manifestations such as cardiac involvement (p=0.027) and pulmonary arterial hypertension (p=0.036). Increased MCP-1 or IFNα serum levels were associated with lung fibrosis (p=0.019 and 0.048, respectively). Digital ulcers including digital loss were associated with increased serum levels of IFNα (p=0.029).
Conclusion An activated type I IFN system previously seen in several other systemic autoimmune diseases is also present in SSc and may contribute to the vascular pathology and affect the profibrotic process.
Statistics from Altmetric.com
Introduction
Systemic sclerosis (SSc) is a rare multisystem autoimmune disease classified either as diffuse (dcSSc) or limited cutaneous (lcSSc), of which the former is more severe with extensive skin and internal organ involvement.1 2 The aetiology of SSc is largely unknown, but gene expression profiles of peripheral blood leucocytes from patients with early SSc demonstrated an increased expression of type I interferon (IFN)-inducible genes—that is, an IFN signature, besides genes involved in targeting blood leucocytes to the endothelium.3 4 Many different autoimmune rheumatic diseases share this IFN signature with SSc, indicating that continuing production of type I IFN is central in autoimmune processes.5,–,9 An increased expression of several chemokines has also been reported in SSc and is of importance for recruitment and activation of inflammatory cells.10 11
We have previously shown that IFNα can be produced by plasmacytoid dendritic cells (pDCs) activated by interferogenic immune complexes (ICs) containing autoantibodies and DNA and/or RNA.12,–,15 It has been reported that SSc sera containing anti-topoisomerase (Topo)-I autoantibodies can form interferogenic ICs, and this ability was associated with lung manifestations in patients.16 In this study, we aimed to clarify further the role of the type I IFN system activation in SSc and to correlate this with clinical manifestations. In an unselected group of patients with SSc we therefore investigated the interferogenic capacity of sera and serum levels of IFNα and the interferon-inducible protein-10 (IP-10)/chemokine (C-X-C motif) ligand 10, monocyte chemoattractant protein-1 (MCP-1)/(C-C motif) ligand-2 (CCL-2), macrophage inflammatory protein-1α (MIP-1α)/CCL-3 and RANTES/CCL-5.
Patients and methods
Patients and controls
A total of 70 patients, fulfilling the 1980 preliminary American College of Rheumatology (ACR) criteria for definite SSc17 were included in the study (table 1).
Demographic characteristics, antinuclear antibody (ANA) pattern and the presence of specific autoantibodies in the patients with systemic sclerosis (SSc) (n=70)*
Gastrointestinal involvement was defined as oesophageal dysmotility, dysphagia, heartburn, constipation, diarrhoea or fecal incontinence. Lung fibrosis was diagnosed by high-resolution CT and/or chest x-ray examination. Cardiac involvement was defined as present or previous cardiac infarction, heart failure, pleuritis or arrhythmia. Presence of digital loss (radiographic evidence of acrolysis or surgical amputation due to necrotic ulcers), pulmonary arterial hypertension (PAH), previous scleroderma renal crisis18 or episodes of peripheral thrombosis were recorded.
Present drugs included proton pump inhibitors (44/70 patients), angiotensin-converting enzyme inhibitors (36/70), calcium channel blockers (24/70), corticosteroids (26/70), non-steroidal anti-inflammatory drugs (22/70) and immunosuppressive drugs (17/70). Serum samples from 30 age- and sex-matched healthy blood donors were used as controls.
Autoantibody determinations
Patient sera were analysed for antinuclear antibodies (ANA) by indirect immunofluorescence using HEp-2 cells and by Crithidia luciliae (Immunoconsept, Sacramento, California, USA) for anti-dsDNA antibodies.12 Patient and control sera were analysed for antibodies to SmB, D, ribonucleoprotein (RNP)-70K, RNP-A, RNP-C, Sjögren syndrome antigen (SSA) (Ro60 and Ro52), SSB, centromere B (CENP-B), Topo-I, Jo-1, ribosomal P antigen and histones by using the Inno-Lia ANA Update line blot assay (Innogenetics, Ghent, Belgium).12
Cell culture conditions and IFNα inducers
Peripheral blood mononuclear cells (PBMCs), pDCs and monocytes were prepared from healthy blood donor buffy coats and cultivated in 96-well plates14; see details in online supplementary material.
Necrotic and apoptotic cell material from monocytic U937 cells13 was used in the cell cultures at a final concentration of 10% and 25%, respectively. The patient or control sera were used in final concentrations of 10%, 1%, 0.1% or 0.01%, with or without necrotic or apoptotic material. Sera were defined as IFNα inducing when IFNα production was higher than the mean IFNα production +2SD induced by control sera together with necrotic (>15 U/ml) or apoptotic (>10 U/ml) material. As a control IFNα inducer, UV-inactivated herpes simplex virus type I was used.13. IgG from six patients with SSc and two healthy control sera were purified as described previously.13
Inhibitors of IFNα production
The necrotic cell material as well as the control IFNα inducers poly I:C (Sigma-Aldrich, St Louis, Missouri, USA), pcDNA3 plasmid and herpes simplex virus were treated with DNase-free RNase A (8 µg/ml; AB-Gene, Surrey, UK) or RNase-free DNase (20 Kunitz units/ml; Qiagen, Hilden, Germany) as described.13 The inducers were incubated with the RNase, DNase or medium only, for 3 h at 37°C before addition of IgG or the transfection agent Lipofectin (Invitrogen, San Diego, California, USA). The anti-FcγRII monoclonal antibody (IV.3: HB-217, ATCC, Rockville, Maryland, USA) prepared as described previously13 and the IgG2b isotype control (BD Biosciences, San Jose, California, USA) were used at indicated concentrations in the induction cultures with PBMCs or pDCs. Chloroquine phosphate (5 µg/ml: Sigma-Aldrich) was used to inhibit endocytosis.
Immunoassay of IFNα
IFNα levels in supernatants were measured by a dissociation-enhanced lanthanide fluoroimmunoassay (DELFIA), that detects most IFNα subtypes but not IFNα2b (detection level ≥2 U/ml).19 To detect IFNα in serum, a more sensitive DELFIA was performed (detection level ≥0.5 U/ml).20
Multiplex analysis of chemokines in patient and control sera
The levels of IP-10, MCP-1, MIP-1α and RANTES were analysed in serum samples by 4-Plex Bio-Plex Multiplex Cytokine Assay (Bio-Rad, Hercules, California, USA). Increased chemokine levels were defined as mean values for the control sera +2SD.
Statistics
Data were analysed using SPSS software (Chicago, Illinois, USA). For analysis of differences between groups, Kruskal–Wallis with the Dunn post hoc test or the Mann–Whitney U test were used. Correlations were calculated using the Spearman rank correlation test. p Values ≤0.05 were considered significant.
Results
Patient characteristics
The patient characteristics are shown in table 1. A total of 55/70 patients were ANA positive. Specific autoantibodies were detected in 43/70 patient sera and in none of the controls. In 10 patients several autoantibody specificities were present: CENP-B and SSA (four patients), CENP-B, SSA and RNP (one patient), SSA and RNP (two patients), SSA and Jo-1 (one patient) and RNP combined with anti-Topo-I in two patients. None had antihistone antibodies and one patient had anti-dsDNA antibodies.
IFNα-inducing ability of the SSc sera combined with apoptotic or necrotic cell material
We initially examined the ability of SSc sera to induce IFNα production in normal PBMCs in combination with either apoptotic or necrotic cell material. We found that 33 and 23 of 70 patient sera at one or more of the tested serum concentrations induced IFNα when combined with necrotic (314±478 U/ml, mean±SD) or apoptotic cell material (399±574 U/ml, mean±SD), which was significantly higher than sera from healthy individuals (figure 1A). All but one of the IFNα-inducing sera induced the highest IFNα levels when 0.1% or 1% SSc serum were used with necrotic or apoptotic material. There was a correlation between the IFNα levels induced by patient sera combined with necrotic and apoptotic cell material (Spearman r=0.537). No IFNα was induced by sera, apoptotic or necrotic cell material alone (not shown).
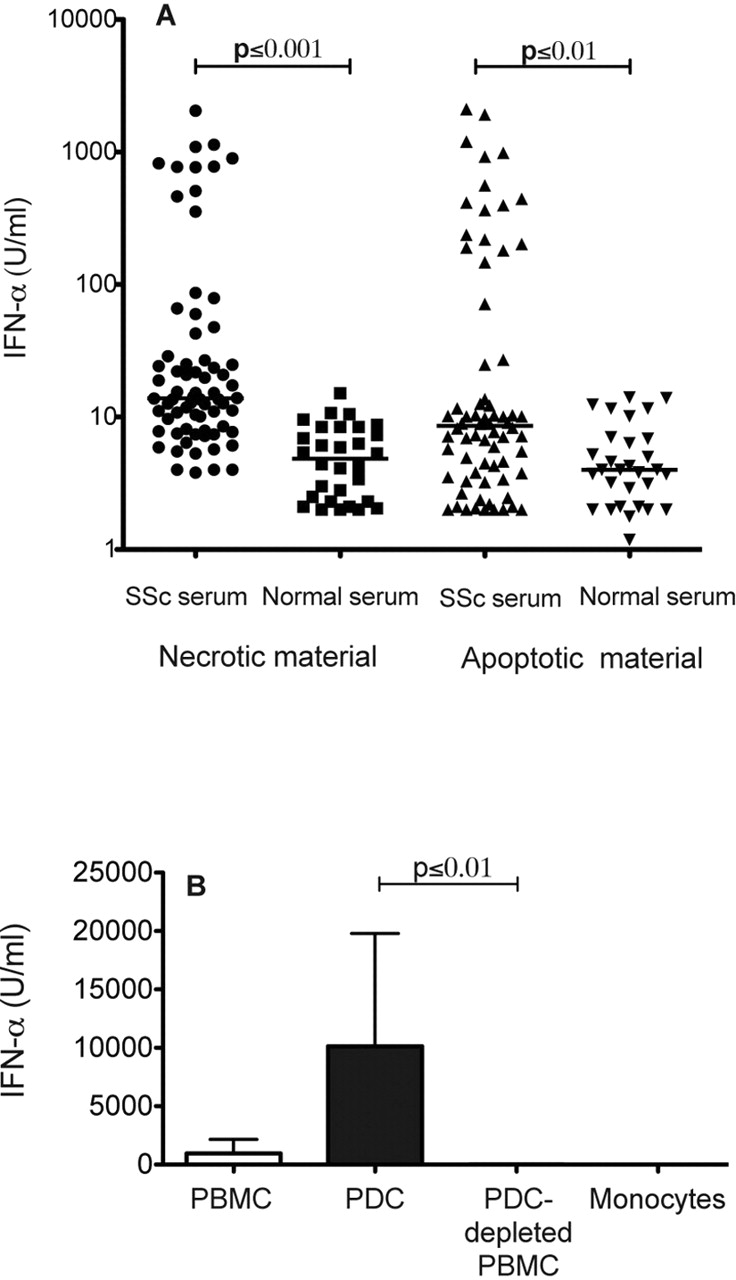
Interferon α (IFNα) production induced by sera from patients with systemic sclerosis (SSc). (A) Peripheral blood mononuclear cells (PBMCs) from healthy blood donors were stimulated by necrotic or apoptotic cell material combined with serum from patients with SSc (n=70) or healthy individuals (n=30). Each data point represents the maximal IFNα level obtained with one of the tested serum concentrations (0.001%, 0.01%, 0.1% or 1%). The horizontal lines show the median values for each group of sera. The experiment was repeated four times with different healthy blood donor PBMCs with similar results. IFNα levels in cell culture supernatants were measured after 20 h by an immunoassay. (B) IFNα production by PBMCs, plasmacytoid dendritic cells (pDCs), pDC-depleted PBMCs or monocytes from healthy blood donors, stimulated by necrotic cell material and sera from patients with SSc. An optimal serum concentration (see A) was used for each SSc serum. Values are mean IFNα levels and standard deviation induced by five SSc sera. One of three experiments with similar results is shown. No IFNα could be detected when sera from healthy controls were used (not shown). IFNα levels in cell culture supernatants were measured after 20 h by an immunoassay.
pDCs are responsible for IFNα production stimulated by SSc-associated IC
We then investigated whether pDCs are responsible for the SSc serum-induced IFNα production. Purified pDCs produced higher amounts of IFNα upon stimulation with SSc sera and necrotic material than PBMCs, and pDC-depleted PBMCs or purified monocytes were unable to produce any IFNα (figure 1B). We then analysed whether immunoglobulins are required for the IFNα production. SSc IgG combined with necrotic cell material-induced IFNα in a concentration-dependent manner (figure 2), but not alone (not shown). The IFNα induction by SSc-associated ICs was sensitive to RNase treatment, but was less affected by DNase (figure 2). Furthermore, the IFNα production by pDCs stimulated with SSc sera and either necrotic or apoptotic material was strongly decreased by antibodies against FcγRII (figure 3A) or by blocking the acidification of endosomes with chloroquine (figure 3B,C). We conclude that pDCs are responsible for the IFNα production triggered by RNA-containing, SSc-associated ICs via an FcγRII- and endosome-dependent pathway.
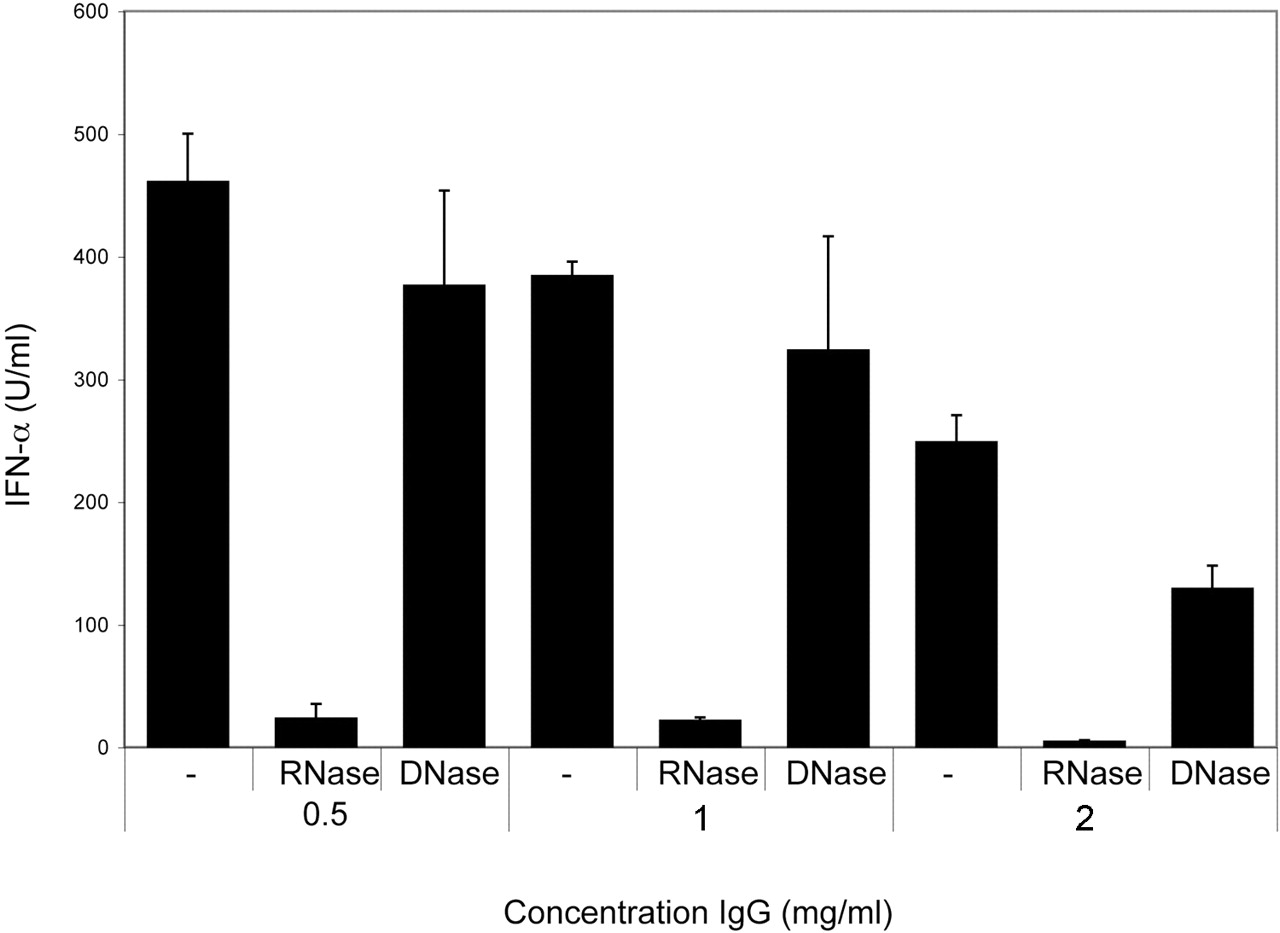
Interferon α (IFNα) production induced by necrotic material and IgG from patients with systemic sclerosis (SSc) requires RNA. Peripheral blood mononuclear cells from healthy blood donors were stimulated with indicated concentrations of SSc IgG and necrotic cell material. The necrotic cell material was treated with RNase or DNase, or left untreated. IFNα levels in cell culture supernatants were measured after 20 h by an immunoassay. The values are means and standard deviations of triplicate wells. Similar results were obtained when several IgG preparations with different autoantibody specificities were used. One of two experiments with similar results using SSc IgG containing autoantibodies to RNP and topoisomerase-I is shown.

Interferon α (IFNα) production in plasmacytoid dendritic cells (pDCs) induced by sera from patients with systemic sclerosis (SSc) is mediated through FcγRII and endosomes. pDCs from healthy blood donors were stimulated with SSc sera and (A, B) apoptotic or (A, C) necrotic cell material. (A) Indicated concentrations of monoclonal antibodies to FcγRII or IgG2b isotype control antibody were added to the induction cultures. (B, C) pDCs were stimulated with SSc sera with different specificities, systemic lupus erythematosus serum or UV-inactivated herpes simplex virus in the presence or absence of chloroquine phosphate (5 µg/ml) and (B) apoptotic or (C) necrotic cell material. IFNα levels in cell culture supernatants were measured after 20 h by an immunoassay. Values are means and standard deviations of at least two different pDC donors in each panel.
Correlation of IFNα induction with the ANA pattern and the presence of specific autoantibodies
Next, we asked if there was any correlation between different ANA patterns or the presence of specific autoantibodies and ability to induce IFNα production. There was no significant difference in the IFNα levels induced in PBMCs by the ANA-negative (n=15) or ANA-positive sera (n=55), combined with either necrotic (117±276 vs 162±381 U/ml; mean±SD) or apoptotic material (98±149 vs 171±441 U/ml; mean±SD). No ANA pattern was associated with significantly higher IFNα-inducing ability of the sera (results not shown).
Sera containing antibodies with multiple specificities induced more IFNα than sera with a single autoantibody specificity. Most sera with multiple autoantibody specificities containing autoantibodies to RNP and/or SSA induced high levels of IFNα (figure 4). A low IFNα-inducing capacity could be found in a proportion of sera containing only autoantibodies to Topo-I or CENP-B, and these sera had no significantly increased IFNα-inducing capacity compared with patient sera lacking these antibodies.

Interferon α (IFNα)-inducing capacity of different autoantibody specificities. Sera from patients with systemic sclerosis (SSc; n=70) or sera from healthy controls (HC; n=30) were combined with (A) necrotic or (B) apoptotic cell material, and added to cultures with peripheral blood mononuclear cells from healthy blood donors. The bars show IFNα levels induced by all sera; HC sera, SSc sera without any detectable autoantibodies (Auto-Ab neg) or SSc sera with a single autoantibody specificity (single) as indicated or with several different autoantibody specificities (multiple). The numbers under the bars are the number of IFNα-inducing sera (inducing more than the mean IFNα production +2SD induced by control sera together with necrotic or apoptotic material) followed by the total number of sera in each category. IFNα levels were measured after 20 h by an immunoassay. Values are means and standard deviations. One of three experiments with similar results is shown.
However, when the 11 sera that induced >200 U/ml of IFNα in combination with necrotic material were analysed in more detail, we found an association with high IFNα-inducing capacity and the presence of ANA with a speckled pattern (p=0.044), anti-RNP (p=0.001) or anti-SSA (p=0.001). The IFNα-inducing capacity of the 16 sera inducing >50 U/ml IFNα with apoptotic material was associated with speckled ANA pattern (p=0.042) and anti-RNP (p=0.001) or anti-SSA (p=0.043).
In vivo production of IFNα, IP-10, MCP-1, MIP-1α and RANTES in patients with SSc
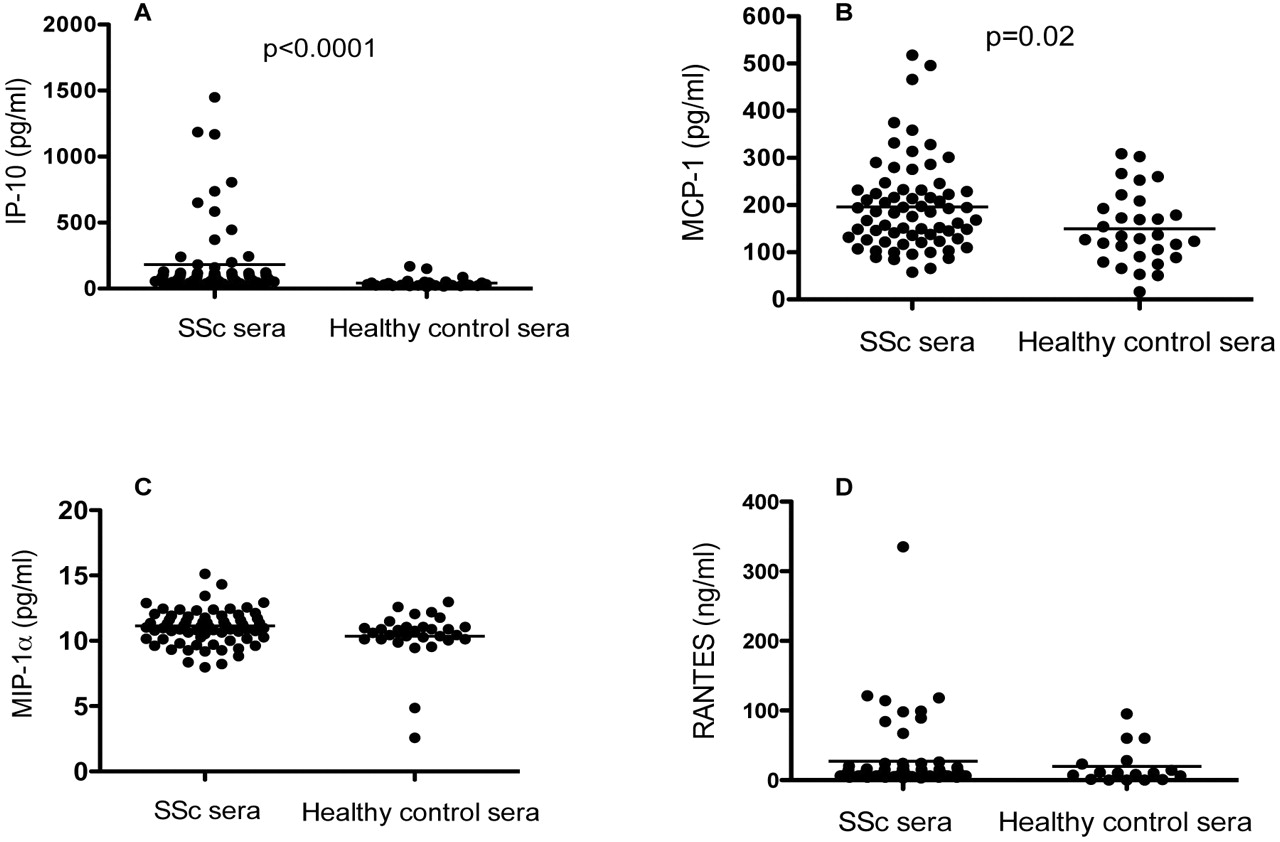
We then explored whether the patients with SSc had any signs of an in vivo activated type I IFN system and if this correlated with some chemokines of importance for the pathogenesis of SSc. We detected IFNα in 7/70 of the patient sera (range 0.5–18 U/ml) (not shown), and 19/69 of the patient sera had increased IP-10 levels (≥118 pg/ml) (figure 5). There was a correlation between IFNα and IP-10 serum levels (Spearman r=0.347, p=0.03). A total of nine of 69 patients had elevated levels of MCP-1 (≥299 pg/ml: p=0.02), which correlated with IP-10 levels (Spearman r=0.309, p=0.009) in the patient sera. Increased levels of MIP-1α (≥14 pg/ml) and RANTES (≥143 ng/ml) were only found in two and one patient samples, respectively. No correlation was found between the serum levels of the measured chemokines and IFNα-inducing ability of the SSc sera.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Serum levels of (A) interferon-inducible protein-10 (IP-10), (B) monocyte chemotactic protein-1 (MCP-1), (C) macrophage inflammatory protein-1α (MIP-1α) and (D) RANTES in patients with systemic sclerosis (n=70) or in healthy controls (n=30) measured by a multiplex bead immunoassay. The horizontal lines show the median values for each group.
Correlation of circulating cytokines with the ANA pattern and specific autoantibodies
We found significantly higher levels of both IP-10 (210 vs 71 ng/ml, p=0.020) and MCP-1 (210 vs 144 ng/ml, p=0.018) in ANA-positive compared with ANA-negative sera. There was a weak association between MCP-1 levels and speckled ANA pattern (p=0.05), while no associations could be found for the other ANA patterns. Antibodies to SSA were associated with increased serum levels of IFNα (p≤0.001) and IP-10 (p=0.011). There was a negative association with autoantibodies to CENP-B and serum IFNα levels (p=0.049), while no other associations could be found between the presence of other specific autoantibodies and the measured cytokines.
Associations between clinical manifestations and the activated type I IFN system
Next we examined possible associations between different disease manifestations and IFNα induction, serum levels of IFNα or the chemokines. Increased levels of IP-10 were associated with cardiac involvement (p=0.027), signs of PAH (p=0.036), a history of arterial or venous thrombosis (p=0.037) or erythrocyte sedimentation rate (ESR) (p=0.002). Increased serum IFNα levels were associated with a history of digital ulcers (p=0.029) and although digital loss was present only in five patients, four of them had increased levels of serum IP-10 compared with one out of 64 without digital loss (Fisher's exact test; p=0.022). Lung fibrosis was associated with increased serum levels of MCP-1 (p=0.019) and IFNα (p=0.048), but not with IFN induction either with apoptotic or necrotic cell material. A history of scleroderma renal crisis was only present in two patients, but both sera induced high IFNα production with necrotic or apoptotic material and also had increased serum levels of IP-10 (see also online supplementary table 1). No association was found either between IFNα induction or serum cytokine levels and other disease manifestations.
We further explored whether ESR as a marker for disease activity might be associated with cytokine production in vivo. We found that serum levels of IFNα (Spearman r=0.39, p=0.001), and the chemokines IP-10 (Spearman r=0.38, p=0.002), MCP-1 (Spearman r=0.25, p=0.045) and MIP-1α (Spearman r=0.46, p<0.001) were correlated with ESR.
We also analysed the sera inducing high levels of IFNα separately and found that IFNα-inducing capacity (>200 U/ml) in combination with necrotic material was associated with ESR (p=0.022) and digital loss (p=0.025) while no such association could be found for apoptotic material.
Discussion
In this study, we observed that sera from an unselected group of patients with SSc together with necrotic or apoptotic material induced IFNα production in pDCs, apparently activated by ICs formed by SSc autoantibodies. Sera from patients with lcSSc or dcSSc were equally effective in triggering IFNα production, demonstrating that the formation of interferogenic ICs is not limited to one of the disease subsets. This finding is corroborated by the observation that patients with lcSSc or dcSSc both display an IFNα signature,4 and that monocytes from patients with lcSSc or dcSSc show no difference in their expression of type I IFN regulated Siglec-1.21 Consequently, both dcSSc and lcSSc can be included in the large group of systemic auto- immune diseases with an IFNα signature that can be caused by the presence of interferogenic ICs that trigger pDCs to a continuous IFNα production.
In line with observations in other diseases, only a proportion of patient sera had the capacity to form interferogenic ICs,12 14 which may be due to several reasons. One possibility is that the type I IFN system activation is most important in the early phases of SSc when immune activation and inflammation is more pronounced than in the later stages of the disease. This explanation is supported by the observations that the highest IFNα production was associated with high ESR. Furthermore, earlier studies showed that circulating IFNα inducers are more common during flares in lupus patients22 23 and that the IFNα signature is more pronounced in early disease.24
When correlating the IFNα production with the presence of different autoantibodies in SSc sera, we found a clear association between induction of high levels of IFNα and autoanti- bodies to RNA-binding proteins—that is, anti-RNP and SSA. Such autoantibodies are frequently present in a number of rheumatic diseases with an IFNα signature5 7 25 and correlate with serum IFNα activity in systemic lupus erythematosus.26 In this study, the interferogenic capacity of formed ICs was also dependent on RNA and was blocked by antibodies to FcγRII as well as by chloroquine. Our results therefore suggest that autoantibodies to RNA-binding proteins are the most important in the generation of interferogenic ICs also in SSc, and indicate that TLR7 activation is a common pathway by which the IFNα signature is expressed in systemic autoimmune rheumatic diseases.
In contrast to a previous study, we could not show that anti-Topo-I antibodies form strong interferogenic ICs.16 The reason for the discrepancy is unclear, but the previous study did not exclude the possibility that other autoantibodies present in investigated sera contributed to the interferogenic activity. We also observed that sera with multiple specificities were most potent in generating interferogenic ICs. Furthermore, in four sera no autoantibodies could be demonstrated either by indirect immunofluorescense for ANA or by the line blot Inno-Lia assay but still induced IFNα production, indicating that the type I IFN production in some patients with SSc is not dependent on ICs or due to interferogenic ICs containing autoantibodies with specificities not investigated in this study. A mechanism for IFNα production not dependent on ICs has been described in psoriasis, where the antimicrobial protein LL37 binds nucleic acid and activates pDCs.27 Thus, similar mechanisms for IFNα production may exist in some patients with SSc.
Few patients had detectable serum IFNα levels and only 28% of the individuals displayed increased IP-10 levels which can be induced by IFNα. However, we observed that increased IP-10 levels were associated with cardiac involvement, signs of PAH, elevated ESR, and although only present in few patients, with digital loss and renal crises. Even though we could not find any associations between the IFNα-inducing capacity and clinical manifestations for the whole group, we found that the high IFNα production (>200 U/ml) with necrotic material was associated with digital loss and increased ESR. We did not correct for multiple testing and the results must be interpreted cautiously, but the observation that most clinical associations were related to vascular manifestations is interesting. The results in our opinion strengthen the view that type I IFN may be important in the development of vasculopathy in SSc. Previous studies in SSc have shown that IFNα mRNA is predominantly expressed in vascular and perivascular cells,4 and that upregulation of myxovirus-resistance protein A28 or an increased expression of IFN genes29 is associated with ischaemic digital ulcers. IFNα has antiangiogenic effects,30 and can impair endothelial cell differentiation in lupus.31 Consequently, activation of the type I IFN system in SSc may affect the microvascular system, and the exact mechanisms by which IFNα contributes to the vascular injury is important to clarify. In addition, there was an association between MCP-1 and IFNα and pulmonary fibrosis, which underscore the importance of the type I IFN system in the disease process.
In summary, a proportion of patients with SSc have autoantibodies that generate ICs with the capacity to activate pDCs to IFNα production in vitro. This observation may at least partially explain the observed IFNα signature in patients with both lcSSc and dcSSc and also the increased serum levels of type I IFN-induced molecules. The association between the latter and clinical manifestations of vasculopathy is intriguing and may explain why many rheumatic diseases characterised by an increased frequency of cardiovascular diseases also have signs of an activated type I IFN system. Taken together, our observations may explain why administration of IFNα to patients with SSc can contribute to progression in the disease process,32 and support the view that the type I IFN pathway is a possible therapeutic target.
Acknowledgments
The authors thank Ms Anne Trönnberg for excellent technical assistance and Ms Rezvan Kiani for collecting the blood samples.
References
Supplementary materials
Web Only Data ard.2009.121400
Files in this Data Supplement:
Footnotes
-
Funding The work was supported by grants from the Dana Foundation, the Swedish Research Council, the Swedish Society of Medicine, the Swedish Rheumatism Association, the Swedish Cancer Foundation, the Uppsala University Hospital Development Foundation, the Agnes and Mac Rudberg Foundation, the Nilsson Foundation, the King Gustaf V 80-year Foundation and COMBINE.
-
Competing interests None.
-
Ethics approval The study was approved by the ethical committee in Uppsala and informed consent was obtained from all patients and controls.