Article Text

Abstract
Objective To analyse Fcγ receptor (FcγR) expression on monocytes and macrophages from rheumatoid arthritis (RA) patients versus healthy controls (HC), and to compare their responses to immune complexes containing RA-specific anti-citrullinated proteins auto antibodies (ACPA).
Methods Monocytes and monocyte-derived macrophages were obtained from the peripheral blood of 34 RA patients and 69 HC. FcγR expression was studied by flow cytometry. Cells were stimulated with ACPA-containing immune complexes, and tumour necrosis factor alpha (TNFα) was assayed in culture supernatants.
Results Variations distinguished RA from HC monocytes, corresponding to a 5% and 6% decrease in the percentages of monocytes expressing FcγRI and FcγRII, respectively, and a 7% increase in the proportion of FcγRIII-positive monocytes. Although in both HC and RA patients macrophage differentiation was accompanied by a dramatic increase in the percentage of FcγRIII-expressing cells (72% vs 74.5%), the parallel decline in the proportion of FcγRI-positive cells was markedly smaller in RA (7% vs 43%). Monocytes and macrophages from patients were responsive to ACPA-containing immune complexes but TNFα production in both cell types neither differed from that observed with the corresponding cells from HC, nor correlated with FcγR expression or clinical or biological data. In RA as in HC, ACPA-containing immune complexes induced secretions of more TNFα in macrophages than in paired monocytes (ninefold). Finally, the proinflammatory potential of ACPA-containing immune complexes was confirmed in CD14-positive monocyte macrophages from the synovial fluid of four RA patients.
Conclusions ACPA-containing immune complexes induce TNFα secretion by blood and synovial fluid-derived macrophages from RA patients, fitting with their probable involvement in RA pathophysiology.
Statistics from Altmetric.com
The efficacy of selective B-cell depletion in rheumatoid arthritis (RA) treatment provides strong evidence for a major involvement of B cells and autoantibodies in this disease. Notably, autoreactivity to citrullinated proteins, manifested by the presence of circulating IgG anti-citrullinated proteins auto antibodies (ACPA), is thought to play a key role.1 2 Indeed, not only are ACPA highly prevalent and very specific for RA, detectable years before disease onset,3 and predictive of future RA development in patients with undifferentiated arthritis,4 but they also have repeatedly been found to be associated with and predictive of an aggressive disease.5 Moreover, the continuous formation of disease-specific immune complexes containing ACPA in the rheumatoid synovium is highly probable. Indeed, in this tissue, not only are ACPA-producing plasma cells present6,–,8 and ACPA concentrated, but also citrullinated proteins are abundant and among them a prominent antigenic target, citrullinated fibrin, has been described.9
Synovial macrophages also play a pivotal role in synovitis. Among the various cell populations of the inflammatory synovial tissue, macrophages are selectively those which number correlates positively with articular pain, inflammation10 and joint destruction,11 and, negatively, with clinical improvement after treatment.12 In addition, macrophages constitute the major synovial tissue source of tumour necrosis factor alpha (TNFα),13 a central proinflammatory cytokine in RA pathogenesis.14 Several mechanisms are thought to be involved in TNFα production including stimulation by interleukin 17, Toll-like receptor engagement and contact with antigen or cytokine-activated T cells.15,–,18 As the isotype distribution of ACPA is compatible with activation of both complement-dependent and Fcγ receptor (FcγR)-dependent inflammatory responses,19 macrophage activation linked to immune complexes containing ACPA is also highly probable. To explore this hypothesis, we developed a human in-vitro model designed to mimic stimulation of the synovial tissue macrophages by immune complexes associating ACPA to synovial deposits of citrullinated fibrin. Macrophages derived from circulating monocytes of healthy blood donors were stimulated by immune complexes generated by immunocapture of the ACPA present in the IgG fraction of a pool of ACPA-positive RA sera, using in-vitro citrullinated human fibrinogen (C-FBG) immobilised on culture wells. TNFα production by the macrophages was evidenced, and a major contribution of the activating FcγRIIa was established, thus demonstrating the capacity of ACPA to mediate a FcγR-dependent proinflammatory response.20 Moreover, the TNFα responses of circulating monocytes to ACPA-containing immune complexes were evaluated and found to be significantly lower than those of the paired macrophages.20
However, among circulating human monocytes, two main populations have been delineated, namely the major FcγRIII-negative and the minor FcγIII-positive populations.21 The latter is considered to be more mature and these cells have been called ‘inflammatory’ monocytes in view of their capacity to produce proinflammatory cytokines and no or little anti-inflammatory mediators.21,–,23 Interestingly, an expansion of this population has been described in inflammatory conditions such as RA,24 Kawasaki disease25 and sarcoidosis.26 Moreover, total circulating monocytes from RA patients with severe disease only treated by non-steroidal anti-inflammatory drugs have been reported constitutively to secrete high amounts of IL-1β, TNFα and prostaglandin E2.27 They were also shown to exhibit increased adhesion and phagocytic activity28 29 compared with monocytes from healthy controls (HC). In addition, in RA patients, a match in the expression of inflammation-related genes such as IL-1α, IL-1β, TNFα, IL-6, GROα, lysozyme and thrombospondin-1 has been observed between that of circulating monocytes and that of macrophages present in the synovial tissue, synovial fluid or rheumatoid nodules.30 Finally, it has been shown that higher levels of bioactive TNFα and matrix metalloproteinase 1 were produced by blood monocyte-derived macrophages from RA patients than by their counterparts from HC, in response to the surrogate immune complexes heat-aggregated gamma globulins (HAGG).31 Therefore, data from the literature suggest that monocytes and monocyte-derived macrophages from RA patients could exhibit disease-associated alterations of their responses to immune complexes, with a possible influence on systemic and articular TNF production. This was investigated in the present study. The FcγR expression profiles and the TNFα responses to ACPA-containing immune complexes were analysed in peripheral blood monocytes and monocyte-derived macrophages from patients with established RA in comparison with cells from HC. Moreover, the assumption that ACPA-containing immune complexes could indeed prompt a TNFα response in articular monocyte-macrophages was validated in a short series of synovial fluid-derived monocyte-macrophages from RA patients.
Methods
RA patients and HC
Blood buffy coats were obtained from 69 healthy adult controls (Établissement Français du Sang Pyrénées-Méditerranée, Toulouse, France). The blood donors ranged in age from 22 to 66 years (median 48 years) and the female to male ratio was 0.31. EDTA venous blood samples were obtained from 34 informed and consenting RA patients attending the Clinical Rheumatology Department of Purpan Hospital. Their demographic, clinical, biological and therapeutic data are shown in table 1. The synovial fluid of two ACPA-negative and two ACPA-positive informed and consenting RA patients was obtained after therapeutic knee or elbow arthrocentesis. All patients fulfilled the 1987 revised American College of Rheumatology criteria for RA.32
Characteristics of RA patients
Flow cytometry
Expression of FcγRI, FcγRII and FcγRIII was analysed by flow cytometry (see supplementary information, available online only, for details on the antibodies used).
Cell culture and stimulation
Monocyte purification from blood or buffy coat samples and differentiation into macrophages were performed as previously described.20 Monocyte-macrophages were purified from synovial fluid samples by direct positive selection of CD14-expressing cells with or without previous elimination of CD15-expressing cells using CD14 and CD15 magnetic beads, respectively (Miltenyi Biotec, Paris, France). Purity, evaluated by May–Grünwald Giemsa coloration, was greater than 90%. Blood-derived or synovial fluid-derived cells were stimulated as previously described.20 Briefly, flat-bottom 96-well plates were coated with C-FBG, and ACPA-containing immune complexes were generated by incubation with an ACPA-positive IgG solution (ACPA+ IgG) prepared from pools of serum samples of patients with RA. After washing, cells were added (50 000/well, three wells/condition) in a serum-free medium (Macrophage-SFM; Gibco, Cergy Pontoise, France) supplemented with sodium pyruvate and antibiotics. Control conditions included culture on C-FBG-coated wells incubated with an ACPA-negative IgG solution (ACPA− IgG) and culture in medium alone or in the presence of lipopolysaccharide. Culture on wells coated with HAGG was also performed. TNFα concentration in 24-h culture supernatants was measured by ELISA (human TNFα ELISA set; eBiosciences, Hatfield, UK), as recommended by the manufacturer.
Statistical analyses
Differences between paired monocyte and macrophage variables were evaluated by Wilcoxon's test. The Mann–Whitney U test was used to compare the variables between RA patients and HC and also between men and women in both groups of patients. Correlations were assessed by Spearman's rank correlation coefficients and significance was determined after Bonferroni correction. For each test, p<0.05 was considered significant.
Results
FcγR expression on monocytes and monocyte-derived macrophages from RA patients
To investigate whether FcγR expression was altered on peripheral blood monocytes and monocyte-derived macrophages from RA patients, their expression of FcγRI, FcγRII and FcγRIII was analysed by flow cytometry in comparison with cells from HC (figure 1).

Comparison of Fcγ receptor (FcγR) expression in monocytes and macrophages from rheumatoid arthritis (RA) patients and from healthy controls (HC). The percentage of positive cells (A) and the mean fluorescence intensity (MFI) (B) were determined by flow cytometry for FcγRI, II and III. Bars represent median values. n, number of donors tested.
Significant variations in the expression of each FcγR were observed at the monocytic stage. Indeed, a small decrease in the proportion of FcγRI-expressing cells was observed in RA (median 90%, range 31–99%) compared with HC (median 95%, range 16–99%; p=0.044) associated with a larger heterogeneity in RA patients (mean±SD: 82±19% in RA vs 91±14% in HC). It was associated with a slight increase in mean fluorescence intensity (MFI) (median 10, range 3–22 in RA vs median 7, range 2–39 in HC; p=0.023). A small decrease in the proportion of FcγRII-expressing cells was also noted in RA (median 91%, range 40–100%) versus HC (median 97%, range 83–100%; p=0.004), associated with a larger heterogeneity in RA (mean±SD: 87±16% in RA vs 96±4% in HC). No changes in MFI were noted. In contrast, FcγRIII expression concerned a higher proportion of monocytes in RA (median 21%, range 5–54%) than in HC (median 14%, range 0–57%; p=0.028), with no significant changes in MFI.
After differentiation into macrophages, the only difference was an important increase in the proportion of cells expressing FcγRI in RA (median 76%, range 25–94%) versus HC (median 51%, range 1–95%; p=0.0004), whereas no significant differences were found for the other receptors and all the MFI were similar in RA patients and HC.
No correlations were found between the expression of each FcγR on RA monocytes or monocyte-derived macrophages (either the percentage of positive cells or MFI) and demographic (age), clinical (disease duration, swollen and tender joints) or biological (erythrocyte sedimentation (ESR), C-reactive protein (CRP), rheumatoid factor (RF) and ACPA titres) data. Similarly, no gender-related differences were detected in the FcγR phenotypes of monocytes and monocyte-derived macrophages, either in HC or in RA patients.
Modulation of FcγR expression during macrophage differentiation in HC and in RA patients
To analyse the modulation of FcγR expression during macrophage differentiation of blood monocytes, we compared the proportion of positive cells for each FcγR in paired monocytes and macrophages, both in RA patients and in HC (table 2). These paired samples constitute a subgroup of the samples presented in figure 1. In RA, macrophage differentiation was accompanied by a highly significant increase in the number of FcγRIII-expressing cells (17.7% positive monocytes vs 92.2% positive macrophages; p=7×10−7), similarly to what occurred in HC (14.1% positive monocytes vs 85.9% positive macrophages; p=2.6×10−14). No differentiation-associated changes in the proportion of FcγRII-positive cells were found either in HC or in RA. Finally, a highly significant decrease in the percentage of FcγRI-positive cells was observed in HC (95.0% positive monocytes vs 52.2% positive macrophages; p=1.2×10−14) whereas the decrease was far less marked in RA patients and did not reach statistical significance (82.9% vs 76.2%; p=0.09).
Modulation of FcγR expression during monocyte differentiation into macrophages*
TNFα secretion of peripheral blood monocytes and monocyte-derived macrophages from RA patients in response to immune complexes
To determine whether, in RA patients, the differentiation of circulating monocytes into macrophages influenced the sensitivity to immune complexes, TNFα secretion was assessed in paired monocytes and macrophages from RA patients in response to iHAGG (19 pairs) and ACPA-containing immune complexes (21 pairs). First, as expected, both types of immune complex induced TNFα secretion by RA cells. Second, overall, as in HC, RA macrophages secreted significantly higher amounts of TNFα than the paired monocytes in response both to iHAGG and to ACPA-containing immune complexes. The detailed results of these assays are provided in supplementary figure S1, available online only.
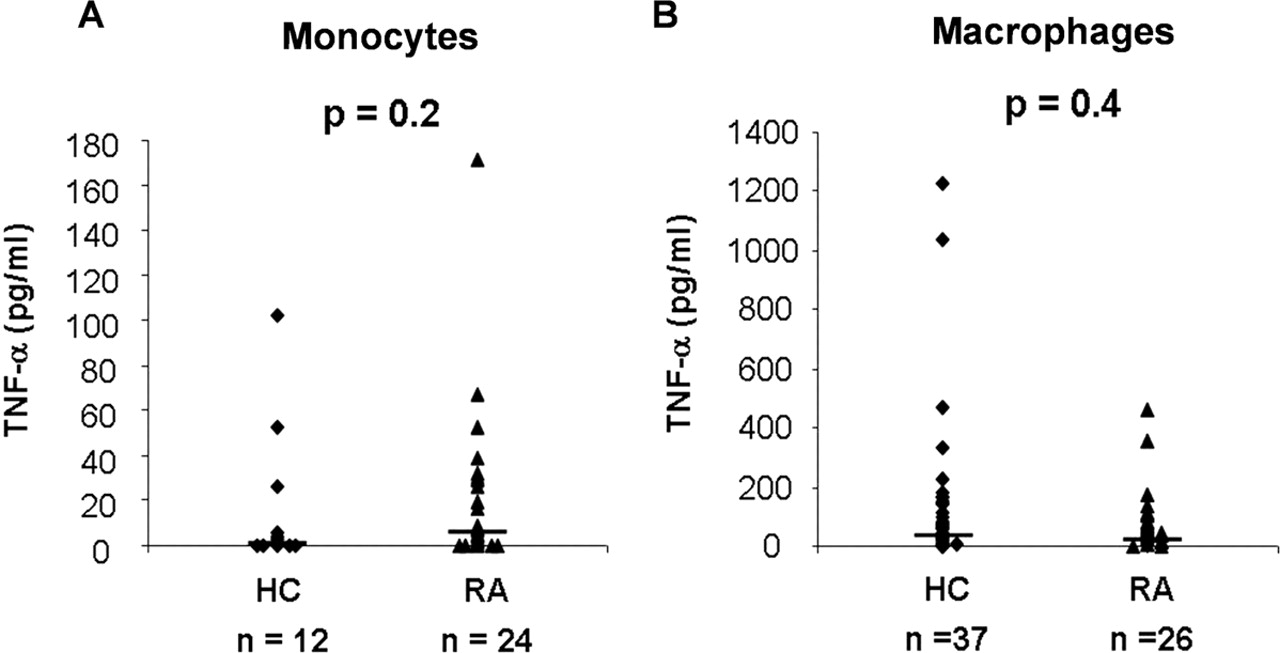
We next assessed whether blood monocytes and monocyte-derived macrophages from RA patients exhibited a particular sensitivity to stimulation by ACPA-containing immune complexes in comparison with cells from HC (figure 2). Monocytes from 12 HC and 24 RA patients showed similar TNFα responses to the immune complexes, with median induced cytokine concentrations of 2 (0–102) and 7 (0–171) pg/ml, respectively (p=0.2; figure 2A). Likewise, the median TNFα concentrations in supernatants of macrophages from 37 HC and 26 RA patients stimulated by ACPA-containing immune complexes were similar, with median TNFα concentrations of 63 (0–1229) and 48 (0–459) pg/ml, respectively (p=0.4; figure 2B).

Tumour necrosis factor alpha (TNFα) responses to anti-citrullinated proteins auto antibodies (ACPA)-containing immune complexes of cells from rheumatoid arthritis (RA) patients and from healthy controls (HC). Monocytes (A) from 12 HC and from 24 RA patients, and macrophages (B) from 37 HC and 26 RA patients were stimulated by ACPA-containing immune complexes. TNFα values (pg/ml) correspond to the difference between the mean TNFα concentrations obtained after culture on citrullinated human fibrinogen (C-FBG)-coated wells incubated with ACPA-positive IgG versus culture on C-FBG-coated wells incubated with ACPA-negative IgG.
No correlations were found between TNFα secretion by RA monocytes or macrophages and the expression of any of the three FcγR on the related cells (percentage of positive cells or MFI) or any clinical (disease duration, tender and swollen joint scores) or biological parameter (ESR, CRP, RF and ACPA titres, evaluated using C-FBG as the immunodetected antigen). Furthermore, there were no significant differences in the sensitivity of monocytes or macrophages to ACPA-containing immune complexes between ACPA-positive and ACPA-negative patients, despite a slight trend towards higher TNFα levels produced by macrophages from ACPA-positive patients (n=20) compared with those from ACPA-negative patients (n=6) (median 51.5 and 31.5 pg/ml, respectively; p=0.52, data not shown).
No differences in TNFα secretion were found between patients using or not using corticosteroids, methotrexate or anti-TNFα biotherapy, considering each medication independently of any other associated drug. Finally, no gender-related differences were detected in the TNFα responses of monocytes and monocyte-derived macrophages from HC and from RA patients, and no correlations were found between these responses and the age of cell donors.
TNFα secretion of monocyte-macrophages from RA synovial fluid in response to ACPA-containing immune complexes
As a further appraisal of the impact on RA pathogenesis of ACPA-induced effector mechanisms, we wanted to verify that ACPA-containing immune complexes could activate monocyte-macrophages isolated from the joints of RA patients. Monocyte-macrophages were isolated by positive selection of the CD14-expressing mononuclear cells of synovial fluid samples from two ACPA-positive and two ACPA-negative RA patients (figure 3). Secretion of TNFα sometimes occurred in the absence of any stimulation (median 2, range 0–135 pg/ml), but irrespective of the ACPA status of patients, it increased when cells were cultured on wells coated with ACPA-containing immune complexes (median increase in TNFα production 26 (18–139) pg/ml), whereas it was not modified after culture on wells in which C-FBG had been incubated with ACPA-negative IgG (median difference in TNFα production 1 (−4 to 8) pg/ml).

{kind=link}
{kind=link}
{kind=link}
Tumour necrosis factor alpha (TNFα) responses to anti-citrullinated proteins auto antibodies (ACPA)-containing immune complexes of monocyte-macrophages from rheumatoid arthritis (RA) synovial fluid. Monocyte-macrophages isolated from the synovial fluid of two ACPA-negative (SF1 and SF4) and two ACPA-positive (SF2 and SF3) RA patients were stimulated by ACPA-containing immune complexes reconstituted by incubating citrullinated human fibrinogen (C-FBG)-coated culture wells with ACPA-positive IgG. TNFα concentrations measured in 24-h culture supernatants were compared with those obtained after culture in medium alone (unstimulated) or on C-FBG-coated wells incubated with ACPA-negative IgG. Bars represent mean levels (error bars are SEM) of triplicate measurements.
Discussion
Whereas our results concerning the expression of FcγR on monocytes from RA patients can be compared with a large set of data from the literature, those concerning FcγR on monocyte-derived macrophages from RA patients can be confronted with a single previously published study.
We found that RA is associated with a significant increase in the number of FcγRI per monocyte, as reflected by the rise in the MFI of staining. Such an increase was also reported in two studies,33 34 but not in two other studies,35 36 and contrasts with the decrease reported in two additional studies.37 38 Our observation that the percentages of FcγRI-positive cells are more heterogeneous and significantly decreased in RA patients compared with HC is unique, and was not reported in the above-mentioned works.
In our series, the MFI for the anti-FcγRII staining was similar in RA patients and in HC, which is in line with results from three other studies.31 36 37 By contrast, as for FcγRI, the percentage of FcγRII-positive cells was more heterogeneous and significantly decreased in RA patients compared with HC. Among the above-mentioned works also studying this receptor,31 35,–,37 only the study by Blom et al31 reported, in a group of 10 RA patients, a percentage of 98% FcγRII-positive cells, no different from that observed in HC cells.
Therefore, concerning FcγRI and FcγRII on circulating monocytes of RA patients, there are wide discrepancies in the literature data concerning their per cell expression. As drugs such as methotrexate,39 infliximab38 or glucocorticoids37 have been reported to influence FcγR expression, the discrepancies could reflect the diversity in the treatments undergone by patients in the various series analysed. In the present study, all patients but two were treated with a disease-modifying antirheumatic drug (DMARD) and/or a biotherapy and/or a glucocorticoid. Therefore, as a result of the small proportion of patients without any treatment or without any DMARD, and due to the treatment diversity, the influence of the various antirheumatic drugs on FcγR expression (either the percentage of positive cells or MFI) could not be formally evaluated.
In contrast, our observation of a higher proportion of circulating monocytes expressing FcγRIII in RA patients in comparison with healthy individuals had previously been reported in various studies,31 35 36 40 despite the treatment diversity. Such an increase in the FcγRIII-positive subpopulation was also reported not only in Kawasaki disease and sarcoidosis, but also in other conditions associated with inflammation, such as atherosclerosis41 and various infectious diseases.42 This suggests that inflammation rather than RA-specific factors contributes to the expression of FcγRIII by a higher proportion of blood monocytes.
More monocyte-derived macrophages from RA patients than their counterparts from HC were found to express FcγRI. This difference results from the significant decrease in the percentage of cells expressing FcγRI following macrophage colony-stimulating factor-induced differentiation, which occurs in HC but not in RA patients. The presence of the FcR γ-chain is required not only for signal transduction through FcγRI and FcγRIII, but also for their surface expression.43 44 In HC, the dampening of FcγRI expression occurring during differentiation into macrophages could be due to a limited availability of the γ-chain induced by the concomitant increase in the expression of FcγRIII. If this is confirmed, it will be interesting to explore whether an increased expression of the FcR γ-chain by monocytes exists in RA, which could account for the lower decrease in FcγRI expression upon differentiation into macrophages.
Concerning FcγRII and FcγRIII, the percentage of positive cells and MFI were not significantly different in the macrophage colony-stimulating factor-induced macrophages from HC and from RA patients. Our observations disagree with those of Blom et al,31 who, using a similar protocol for macrophage differentiation, reported a higher MFI for FcγRII and higher percentages of positive cells for FcγRIII, in a series of 10 RA patients compared with 10 HC. A striking divergence with our study is the absence of patients undergoing anti-TNFα biotherapy in the analysed series. This may partly account for the discrepancy concerning FcγRIII expression because, when confining our analyses to RA patients without anti-TNFα biotherapy, the trend towards an increase in the percentage of FcγRIII-expressing cells already visible in the whole patients series was more pronounced, even though it did not reach statistical significance. However, in that subgroup we did not detect any rise in the MFI for anti-FcγRII staining.
We have previously demonstrated the proinflammatory potential of ACPA-containing immune complexes using monocytes and monocyte-derived macrophages from healthy individuals.20 Here, we extended this demonstration to cells obtained from RA patients.
RA monocytes were responsive to ACPA-containing immune complexes, but their TNFα response was not significantly different from that of HC monocytes, despite their phenotypical differences, notably the increased proportion of FcγRIII-positive ‘inflammatory’ monocytes. However, one should stress that for all three FcγR, modifications were rather modest and the absence of a significant functional consequence at the level of the whole monocyte population might not be surprising on the limited series analysed.
In RA patients as in HC, we noted that activation by iHAGG and by ACPA-containing immune complexes was globally lower in monocytes than in the derived macrophages, except for two patients in whom monocytes were curiously more responsive than macrophages (supplementary figure S1). None of the clinical or phenotypical characteristics of the two patients could explain this paradoxical result. Compared with those from HC, macrophages from RA patients did not exhibit altered TNFα responses to ACPA-containing immune complexes, despite the important increase in the proportion of cells expressing FcγRI. This is compatible with our previous observations suggesting that this receptor contributes minimally to the TNFα response of macrophages to such immune complexes.20 Given the major contribution of FcγRII to the immune complex-induced TNFα response, the lack of significant difference in FcγRII expression between macrophages from RA patients and those from HC is compatible with a lack of difference in their response to immune complexes. However, it would have been interesting to investigate separately the possible alterations in the expression of the activating and the inhibiting forms of FcγRII. However, so far, studies in which the expression of FcγRIIa was specifically examined in RA patients have brought discrepant results, because a decreasing trend of the related MFI was reported by Hepburn et al36 while Wijngaarden et al34 noted a significant increase.
Differences in sensitivity to immune complexes might be found in non-treated RA patients, in comparison with HC, but the set up of such a study is made difficult by the scarce availability of blood from such patients. Only a longitudinal study performed on patients in whom monotherapies would be initiated could allow first to evaluate whether RA cells are hypersensitive to immune complexes before treatment, and then what type of treatment could have an impact on this sensitivity.
Growing evidence suggests that ACPA-positive and ACPA-negative RA correspond to two disease subsets with different genetic predisposition, severity and probably pathophysiological mechanisms.45 We compared FcγR expression and sensitivity to immune complexes between cells from 26 ACPA-positive and eight ACPA-negative RA patients. Even if these results need to be confirmed on a larger series of ACPA-negative patients, the absence of significant differences suggests that FcγR expression and in-vitro sensitivity of blood-derived cells to immune complexes are not key differential factors in these two groups of patients.
In preliminary experiments, we verified that ACPA-containing immune complexes could stimulate TNFα secretion by monocyte-macrophages isolated from the synovial fluid of four RA patients. Observing that ACPA-containing immune complexes induced TNFα secretion allows confirmation of the proinflammatory potential of ACPA in an in-vitro setting that is even more relevant to the disease pathophysiology. Studies in larger series allowing comparisons between synovial fluid monocyte-macrophages from RA and non-RA patients and between blood monocyte-derived macrophages and monocyte-macrophages from the synovial fluid and/or synovial tissue in the same RA patients were definitely beyond the scope of the present study. However, they would undoubtedly bring interesting additional information as they would allow an evaluation of how disease-associated phenotype(s) of articular monocyte-macrophages influence the cytokine response to ACPA.
The in-vitro model we used here tests the effect of solid phase immune complexes as its purpose is to imitate the encounter of macrophages with ACPA having interacted with citrullinated fibrin deposits present in the synovial tissue of RA patients. Obviously, given the presence of soluble ACPA-containing immune complexes in the synovial fluid of RA patients,46 it would also be interesting to evaluate the capacity of such immune complexes to promote macrophage or polymorphonuclear cell activation and therefore to participate in the inflammatory response within this compartment. However, even if of potential interest, notably because some divergences have been reported concerning the functional effects of soluble versus insoluble immune complexes in different cellular models,47 48 a formal comparison between the effect of soluble and that of solid phase ACPA-containing immune complexes appears technically laborious as it requires the availability of purified ACPA and only makes sense if cells are in contact with equal amounts of ACPA when testing the two immune complex categories.
Finally, we demonstrated that ACPA-containing immune complexes induced TNFα secretion in the absence of serum and therefore in a complement-independent manner. However, in an in-vitro assay, Trouw et al49 recently reported that ACPA could also activate classic and alternative complement pathways. Moreover, a recent study demonstrated that the formation of immune complexes containing ACPA of the IgE class derived from the serum of ACPA-positive RA patients at the surface of basophils led to their activation, as reflected by the induction of CD63 expression.50 Taken together, all these data reinforce the hypothesis of a direct involvement of ACPA in RA pathophysiology, through activation of major proinflammatory effector functions, and reinforce the potential relevance of therapeutic approaches aimed at counteracting these effector pathways.
Acknowledgments
The technical assistance of Ms R Llobera, M-F Isaïa, M-P Henry, L Ceccato and C Gohory is gratefully acknowledged. The authors also thank F-E L'Faqihi-Olive and V Duplan-Eche (Plateau technique de Cytométrie, IFR-150, Toulouse) for technical assistance in flow cytometry analyses.
References
Supplementary materials
Web Only Data
Files in this Data Supplement:
Footnotes
LL and CC and GS and MS contributed equally to this work.
-
Funding This study was supported by grants from the Toulouse III University, the ‘CNRS’ and the ‘Fondation Arthritis’.
-
Competing interests None.
-
Patient consent Obtained.
-
Ethics approval The use of blood samples has been approved by the Local Committee for the Protection of Persons (Sud Ouest et Outre-Mer I) and has been declared to the Ministry of Higher Education and Research (file no. DC-2008-264).
-
Provenance and peer review Not commissioned; externally peer reviewed.