Article Text

Abstract
Objectives Interleukin (IL)-17 signalling has been shown to be a key regulator of disease in ankylosing spondylitis (AS) with several IL-17 blockers currently clinically approved. Despite this, the role of IL-17 in bone pathology is poorly understood. This study aimed to investigate IL-17 signalling in the context of pathological bone formation.
Methods A biomimetic human periosteum-derived cell (hPDC) model of osteogenic differentiation was used in combination with recombinant IL-17 cytokines, T-cell supernatants or serum from patients with AS. IL-17A, IL-17F and bimekizumab monoclonal antibodies were used to block IL-17 cytokine action.
Results Recombinant IL-17A and IL-17F are pro-osteogenic with respect to hPDC differentiation. T helper 17 or γδ-T cell supernatants also potently stimulated in vitro bone formation, which was blocked deeper by dual inhibition of IL-17A and IL-17F than by neutralisation of IL-17A or IL-17F individually. Osteogenic blockade may be due to an increase in expression of the Wnt antagonist DKK1. Interestingly, osteocommitment was also induced by serum obtained from patients with AS, which was also abrogated by dual neutralisation of IL-17A and IL-17F.
Conclusions These data show for the first time that IL-17A and IL-17F enhance in vitro osteogenic differentiation and bone formation from hPDCs, inhibition of which may offer an attractive therapeutic strategy to prevent pathological bone formation.
- TCells
- Cytokines
- Ankylosing Spondylitis
- Inflammation
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
INTRODUCTION
Pathological bone formation associated with spondyloarthritis (SpA) is a major cause of structural tissue damage, which results in permanent disability.1 Ankylosing spondylitis (AS) is the most characterised SpA with regards to pathological bone formation, and recent experimental and clinical studies have highlighted a potential role for interleukin (IL)-17A signalling in this process.2 3 Specifically, IL-17A inhibition with secukinumab results in the arrest of radiographic progression in almost 80% of treated patients with AS over 4 years of treatment.3 However, the cellular basis for this observation is largely unknown with conflicting reports on the effect of IL-17A on osteoblast lineage cells. IL-17F is a close homologue to IL-17A and has also been proposed to have a pro-osteogenic capacity in vitro and in vivo4; however, data are limited compared with those for IL-17A. While IL-17A and IL-17F have been implicated in osteoblast biology, a comparative analysis of their ability to modulate osteogenic differentiation in human disease-relevant cells has yet to be reported. Interestingly, IL-17A-producing γδ-T cells have a critical function in periosteal bone formation for fracture repair,5 while the periosteum has also been implicated in pathological bone formation during SpA disease progression.1 6
After fracture, progenitor cells in the periosteum undergo rapid expansion and differentiation into chondrocytes and osteoblasts to facilitate repair through endochondral or direct mechanisms.7 Fracture healing is known to be highly dependent on an initial inflammatory response, with anti-inflammatories having a negative effect on repair.8 Many similarities can be drawn between pathological bone formation and fracture repair, and as such we hypothesise that pathological bone formation is analogous to ‘fracture repair in the absence of fracture’. However, this hypothesis has only recently become testable due to our work on the creation of in vitro systems that faithfully replicate human periosteal biology.
Herein, an in vivo inspired biomimetic human periosteum-derived cell (hPDC) differentiation methodology was used to probe IL-17-mediated outcomes related to periosteal bone formation.9 Additionally, we investigated whether bimekizumab, a humanised monoclonal antibody that potently and selectively neutralises IL-17A and IL-17F,10 could suppress inflammation-driven in vitro bone formation.
MATERIALS AND METHODS
hPDC cultures
hPDCs were isolated from biopsies obtained from patients undergoing orthopaedic surgery11 12 (obtained from Prof. Frank Luyten, KU Leuven, Belgium). Subsequently, hPDCs from a pool of six donors were expanded in growth medium (GM) consisting of high-glucose Dulbecco’s Modified Eagle’s medium (DMEM, Invitrogen, Paisley, UK) supplemented with 10% batch-tested fetal bovine serum and antibiotic–antimycotic solution (Invitrogen) to passage 5. All experiments described herein were performed in pooled hPDCs at passage 6. We have previously published the CD marker profile (CD73+/CD90+/CD105+), tri-linage differentiation capacity (osteogenic/chondrogenic/adipogenic) and in vivo bone-forming capacity (bone ossicle with marrow) of this donor pooled population.11 13
Generation of Th17 and γδT cell supernatants
Peripheral blood mononuclear cells (PBMCs) from healthy donors were stained with anti-human antibodies against CD4 (RPA-T4), CD45RO (UCHL1), CCR6 (R6H1) and CXCR3 (CEW33D) to identify and sort Th17 (T helper 17) cells with the following expression profile: CD4+CD45RO+CCR6+CXCR3−. For γδ T cells, PBMCs were stained with anti-CD3 (UCHT1), γδTCR (B1.1), CXCR3 and CCR6 (R6H1). The resulting cells were sorted using a BD-FACS Aria III flow cytometer. The cells were then stimulated with plate-bound anti-CD3 (OKT3; 1 µg/mL) and soluble anti-CD28 (CD28.2; 1 µg/mL), and the supernatant was harvested after 5 days. The resultant supernatant was analysed by multi-array Meso Scale Discovery (MSD) for interferon γ (IFNγ), IL-10, IL-1β, IL-6 and tumor necrosis factor α (TNFα) (MSD V-Plex pro-inflammatory panel, MSD—K15049D-1), as well as by single ELISA for IL-17A (DY317-05) and IL-17F (DY1335B-05) ELISA (R&D Systems).
In vitro osteogenic assays
hPDCs were seeded at a density of 3000 cells/cm2 in GM, and after 24 hours, the culture media replaced with either GM or the biomimetic growth factor cocktail (GFC). GFC was prepared through supplementation of GM with 10 ng/mL IL-6; 20 ng/mL epidermal growth factor; 10 ng/mL transforming growth factor-β1; 57 µM ascorbate-2-phosphate; 3 mM Ca2+ and 2 mM PO4 - ions, or without IL-6 supplementation (referred to as differentiation medium; DM).9 Experiments involving the stimulation with IL-17 cytokines were conducted through the supplementation of DM media with 50 ng/mL recombinant IL-17A or IL-17F. Alternatively, GM was supplemented with Th17 or γδ-T cell supernatants (1:50 dilution) and pre-incubated with 10 µg/mL high-affinity antibodies to IL-17A, IL-17F or bimekizumab for 1 hour in a shaking water bath at 37°C. DM components were added to the mixture and applied to hPDCs. All cultures were maintained in 5% CO2, with media replaced every other day until termination.
Gene expression analysis
Total RNA was extracted using the RNeasy mini kit (Qiagen, Manchester, UK) as per manufacturer’s instructions and reverse transcribed into complementary DNA (cDNA) using the high-capacity cDNA reverse transcription kit (Applied Biosciences, Bedford, UK). To detect messenger RNA (mRNA) transcripts, primers (exon spanning, Primer3 Plus, NCBI) were premixed with iTaq universal SYBR green supermix (Bio-Rad, Herts, UK) and combined with cDNA in 10 μL reactions. Thermal cycling conditions were as follows: 10 min at 95°C, with 40 cycles of 15 s at 95°C, 30 s at 60°C and 20 s at 72°C, on a Bio-Rad CFX1000 Real-Time System (Bio-Rad). Target gene quantification was achieved using the 2-ΔΔCT method relative to HPRT1 as the housekeeping gene. Primer sequences are listed in online supplementary table 1.
Supplementary file 1
Assessment of matrix mineralisation
Monolayers were washed with phosphate-buffered saline (PBS) and subsequently fixed with 10% neutral buffered formalin for 20 min at room temperature. After several washes with PBS, cell layers were stained with aqueous 1% (w/v) alizarin red solution (Sigma) at pH 4.2 for 10 min at room temperature, before washing with 50% ethanol to remove any unbound stain. Culture wells were scanned using an Epson flatbed scanner (Epson, UK) and photomicrographic image of each culture well was captured to record the distribution of mineral staining. The bound stain was subsequently eluted with 10% cetylpyridinium chloride (Sigma), and the optical density of the resultant solution determined at 595 nm (Tecan Sunrise, Mannedorf, Switzerland).
Human AS patient serum isolation and effect on hPDC osteocommitment
Blood from patients with AS was isolated in BD Vacutainer SST tubes in the rheumatology clinic at the Royal National Orthopaedic Hospital (Stanmore, UK). This study was approved by the NHS Research Ethics Committee (research authority: Yorkshire and the Humber—Leeds West Research Ethics Committee 16/YH/0137). After collection, the blood was centrifuged at 1000 g for 10 min at room temperature. The separated serum was then filtered through a 0.2 μm membrane and stored at −80°C. hPDCs were seeded at 10 000 cells/cm2 in 24-well plates for 24 hours, following which the media was removed and the monolayers were washed with sterile warmed PBS. Subsequently, DMEM containing either 10% healthy human serum (HS) or serum from patients with AS (AS1, AS2, AS3, AS4, AS5 and AS6) with 1% sodium pyruvate and 1% antibiotics/antimycotics was added to the cultures for 48 hours. The monolayers were then lysed for total RNA isolation. A selected cohort of the serum was pre-incubated with 10 µg/mL high-affinity antibodies to IL-17A, IL-17F, bimekizumab or control IgG for 1 hour in a shaking water bath at 37°C. The serum/antibody mix was then applied to hPDCs and analysed as described earlier.
Statistical analysis
Data are expressed as mean±SEM. Statistical significance was determined using either the two-tailed Student’s t-test with Welch’s correction, one-way analysis of variance (ANOVA) or one-way ANOVA with Fisher’s least significant difference (LSD) post hoc corrections applied. Statistical significance is indicated on all graphs as follows: *p<0.05, **p<0.01, ***p<0.001 (n=3). All statistical analysis was performed using GraphPad Prism version 6.0f for windows (GraphPadSoftware, La Jolla, California, USA, www.graphpad.com).
RESULTS
IL-17RA and IL-17RC expression is increased during osteogenic differentiation of hPDCs
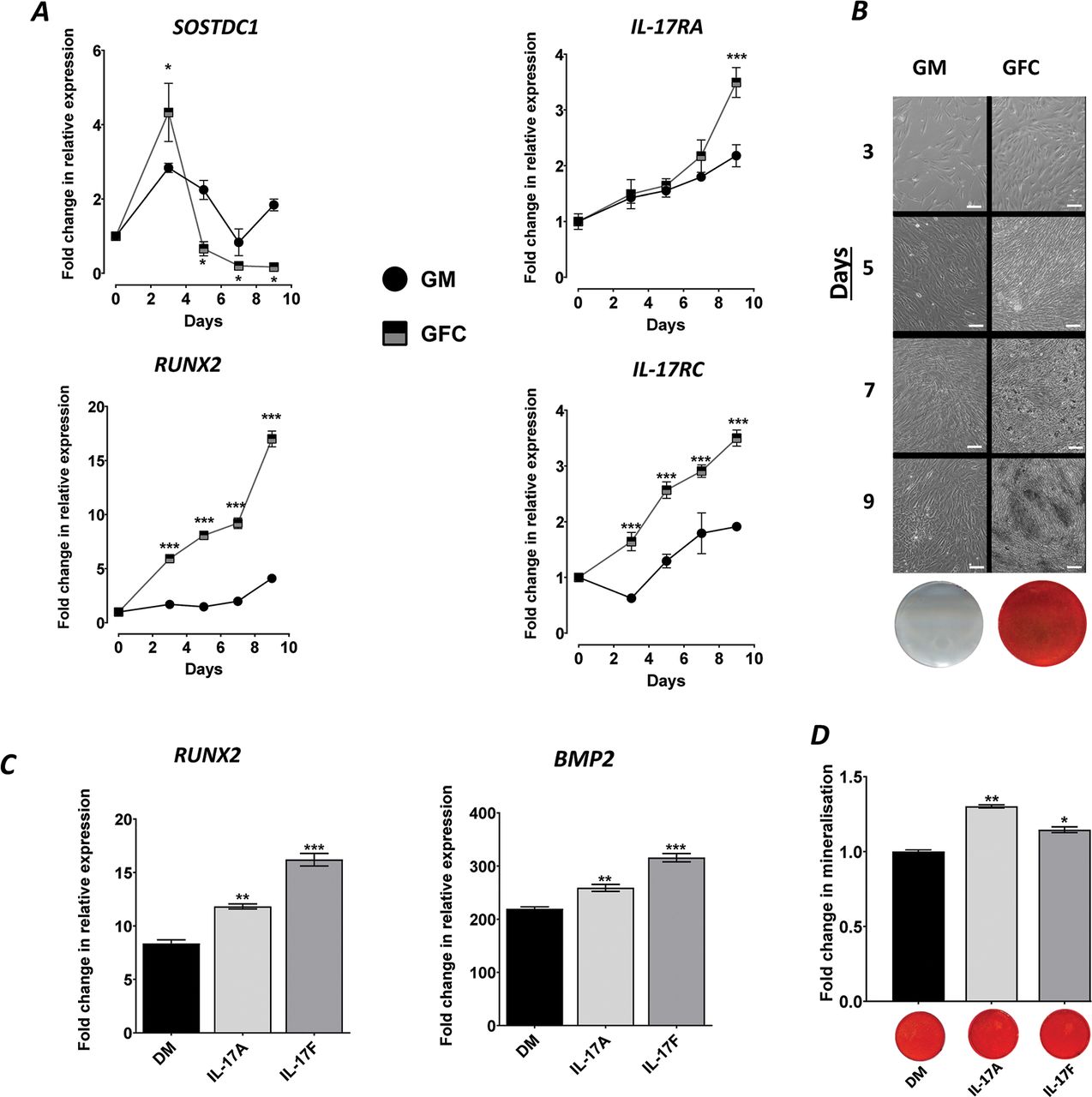
To probe periosteum-associated IL-17 biology, we used an hPDC model that recapitulates in vivo-type bone formation in vitro through the use of a defined osteogenic GFC.9 During differentiation, the expression of the periosteal stem cell marker SOSTDC1 peaked at day 3 and then subsequently decreased over time indicating differentiation away from a ‘stem cell’ phenotype (p<0.05 at all timepoints vs GM). A simultaneous increase in the expression of the osteocommitment marker RUNX2 was observed by day 3, which was approximately fourfold higher than GM by day 9 (p<0.001) (figure 1A). Furthermore, the expression of the IL-17A and IL-17F receptors, IL-17RA and IL-17RC, was increased during osteogenic differentiation compared to GM (p<0.001; figure 1A). In vitro bone formation was confirmed by the potent increase in bone nodules and mineralisation (figure 1B).
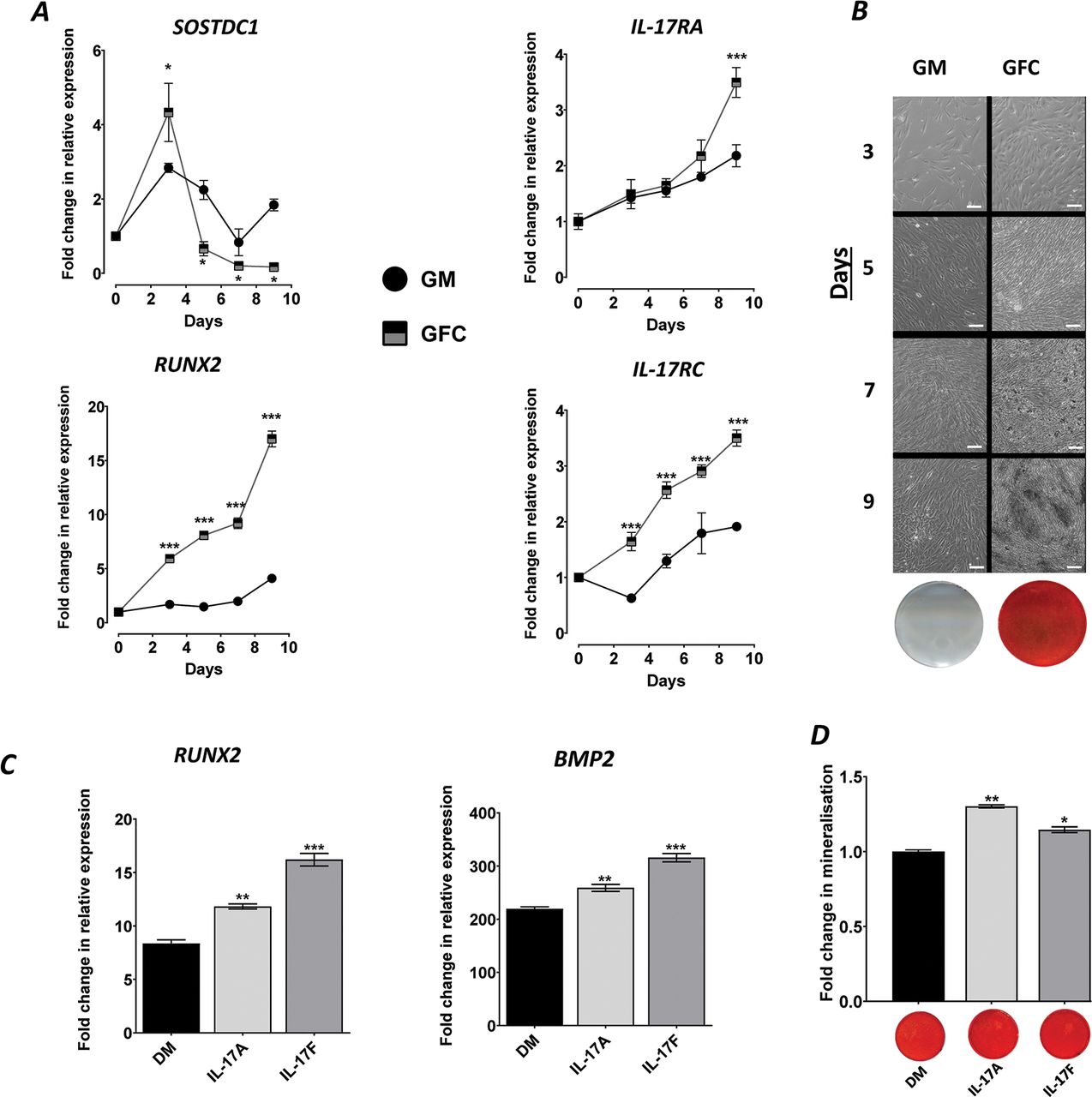
IL-17 is intimately involved with osteogenic differentiation of hPDCs. (A) Expression of SOSDC1, RUNX2, IL-17RA and IL-17RC in hPDCs cultured in GM or GFC. Results are expressed as the mean fold change in expression compared to day 0 GM±SEM (n=3). (B) Representative phase-contrast images of hPDCs during 3, 5, 7 and 9 days of culture under GM or GFC conditions showing bone nodule formation (dark regions from day 7) in GFC, with corresponding day 9 alizarin red–stained monolayer images. (C) Expression of RUNX2 and BMP2 cultured under DM or supplemented with 50 ng/mL rhIL-17A or 50 ng/mL IL-17F for 9 days. Results are expressed as the mean fold change in expression compared to day 0 GM±SEM (D) Quantification of matrix mineralisation in hPDCs cultured in DM or supplemented with 50 ng/mL IL-17A or 50 ng/mL IL-17F for 9 days. Representative images of matrix mineralisation stained with alizarin red are presented below the quantification graph. Results are expressed as the mean fold change in expression compared to day 0 GM±SEM. ***p<0.001; **p<0.01; *p<0.05; comparisons between each condition by one-way ANOVA with Fisher’s LSD post hoc test (n=3). ANOVA, analysis of variance; DM, differentiation medium; GM, growth medium; GFC, growth factor cocktail; hPDCs, human periosteum-derived cells; IL, interleukin.
IL-17A and IL-17F cytokines promote osteogenic differentiation and matrix mineralisation
Due to the minor role that IL-6 plays in hPDC differentiation9 and due to potential for cytokine synergies/interactions, IL-6 was removed from the GFC in all subsequent experiments (referred to as DM). Stimulation with 50 ng/mL IL-17A or IL-17F for 9 days resulted in a significant increase in the expression of RUNX2 (transcriptional regulator of osteogenesis; p<0.01; figure 1C) and matrix mineralisation compared to DM (figure 1D). The expression of BMP2, which has previously been shown to synergise with IL-17 cytokines,4 was also enhanced (p<0.01; figure 1C). Analysis of additional osteogenic markers (SP7, BGLAP, VEGFA and PHOSPHO1) indicated that both cytokines enhance osteogenic differentiation of hPDCs in an approximately equipotent manner (online supplementary figure 1).
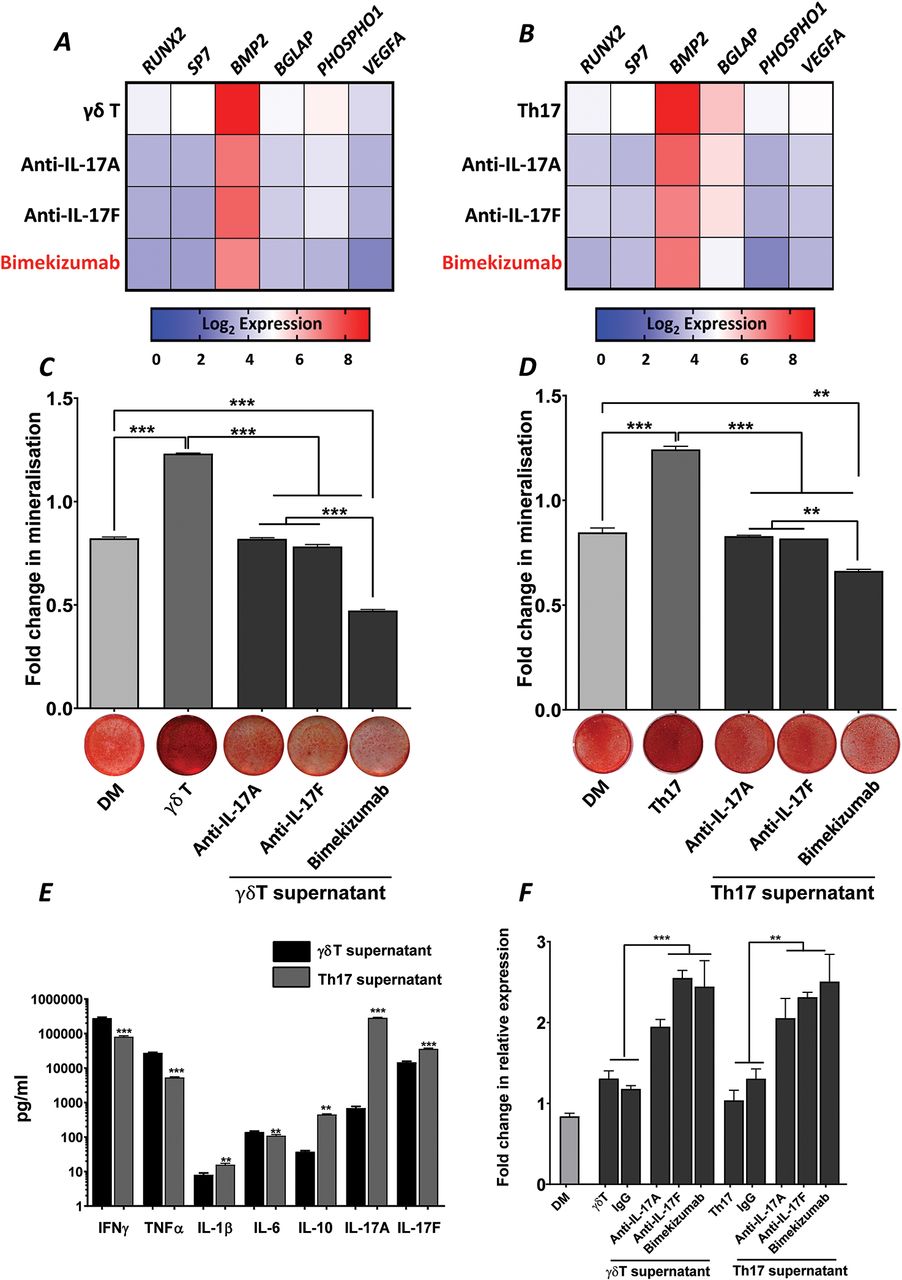
Dual neutralisation of IL-17A and IL-17F demonstrated greater suppression of γδ and Th17 T cell-mediated osteogenic differentiation of hPDCs than blockade of IL-17A or IL-17F alone
We have previously used activated Th17 cell supernatants to mimic the pro-inflammatory milieu and assess the functional contributions of IL-17A and IL-17F in vitro.10 Herein, supernatants from γδT cells, which have previously been shown to contribute to periosteal bone formation, have also been analysed. Exposure of hPDCs to γδT or Th17 cell supernatants in DM resulted in potent increases in all osteogenic markers (online supplementary figure 2) and matrix mineralisation (figure 2C and D). The combined gene expression heat maps in figure 2A and B indicate that the dual neutralisation of IL-17A and IL-17F induces a deeper suppression of osteogenic gene expression than the neutralisation of either cytokine alone. The functional result of this was the suppression of matrix mineralisation to below levels seen with DM (figure 2C and D). Interestingly, when comparing the cytokine composition of the supernatants (figure 2E), an inverse IL-17A/IL-17F ratio was observed, which was in favour of IL-17F for γδT cells. This may explain subtle differences in osteogenic gene expression profile induced by γδ and Th17 cell supernatants (figure 2A and B). Interestingly, a significant (p<0.001) increase in the expression of DKK1 was observed in all samples in response to IL-17 neutralisation (figure 2E).
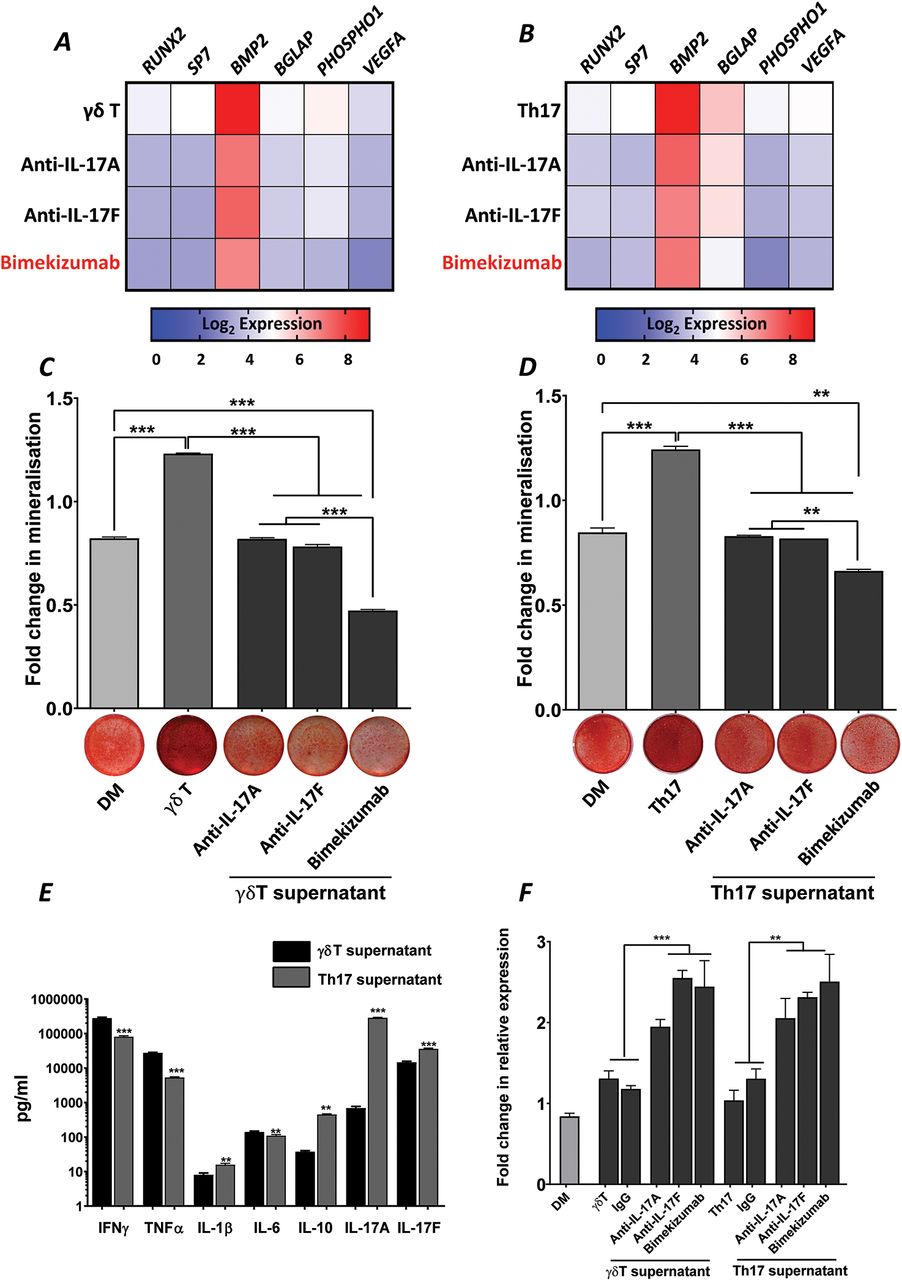
Bimekizumab suppresses γδ T cell and Th17 supernatant-mediated osteogenic differentiation of hPDCs. Heat maps summarising inhibition of osteogenic marker expression in hPDCs stimulated with DM or DM supplemented with (A) γδ T cell supernatant or (B) Th17 cell supernatant (± preincubation with 10 µg/mL anti-IL-17A; anti-IL-17F or bimekizumab) for 9 days. Results are expressed as the Log2 mean fold change in expression compared to growth media control. Quantification of matrix mineralisation after 9 days incubation in (C) γδ T cell supernatant or (D) Th17 cell supernatant (± preincubation with 10 µg/mL anti-IL-17A; anti-IL-17F or bimekizumab). Representative images of matrix mineralisation stained with alizarin red are presented below the quantification graphs. Results are expressed as the mean fold change in expression compared to GM±SEM. ***p<0.001; **p<0.01; *p<0.05; comparisons between each condition by one-way ANOVA with Fisher’s LSD post hoc test (n=3). (E) MSD-multi-array v-plex pro-inflammatory panel expression of IFNγ, TNFα, IL-1β, IL-8, IL-10, IL-17A, IL-17F cytokines from Th17 and γδT cell supernatants. Results are expressed as mean pg/mL ± SEM. ***p<0.001; **p<0.01; comparisons between Th17 and γδT cell supernatants by two-tailed Student’s t-test for each cytokine tested. (F) Expression of DKK1 in hPDCs stimulated with DM or DM supplemented with either γδ T cell or Th17 cell supernatant (± preincubation with 10 µg/mL anti-IL-17A; anti-IL-17F or bimekizumab) for 9 days. Results are expressed as the mean fold change in expression compared to day 0 GM ± SEM. ***p<0.001; **p<0.01; *p<0.05; comparisons between each condition by one-way ANOVA with Fisher’s LSD post hoc test (n=3). ANOVA, analysis of variance; DM, differentiation medium; GM, growth medium; hPDCs, human periosteum-derived cells; IL, interleukin; LSD, least significant difference; MSD, Meso Scale Discovery; Th17, T helper 17; TNF, tumor necrosis factor.
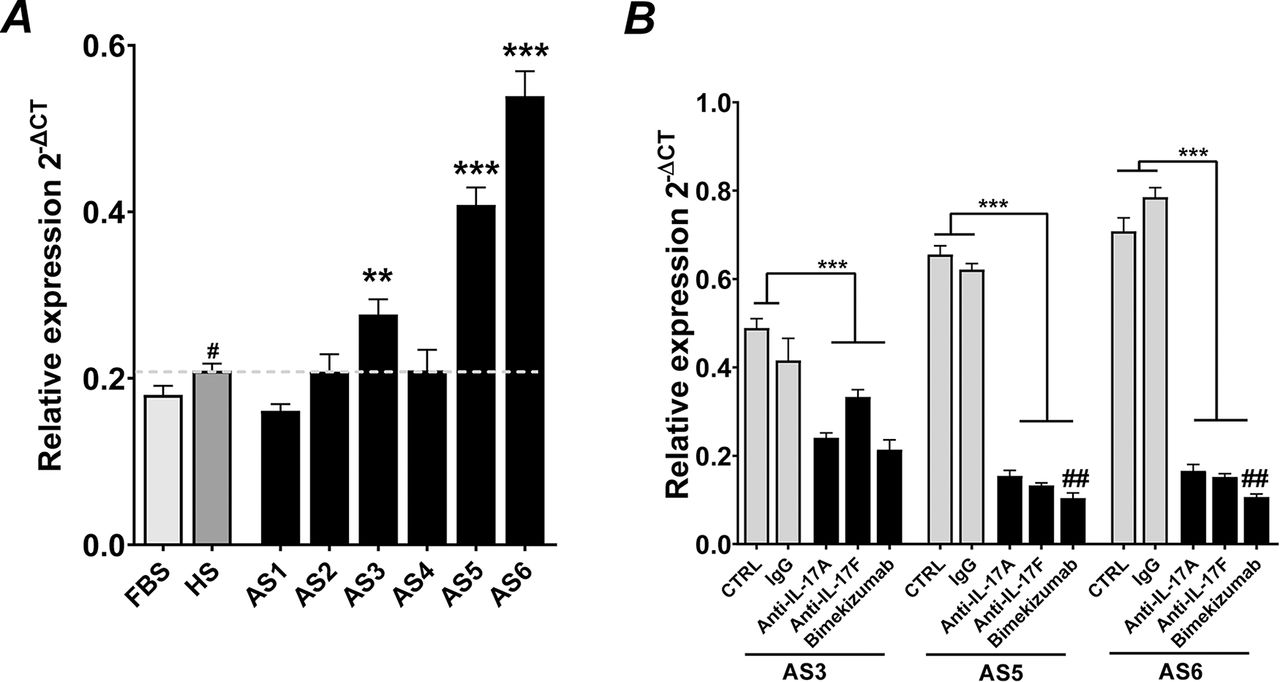
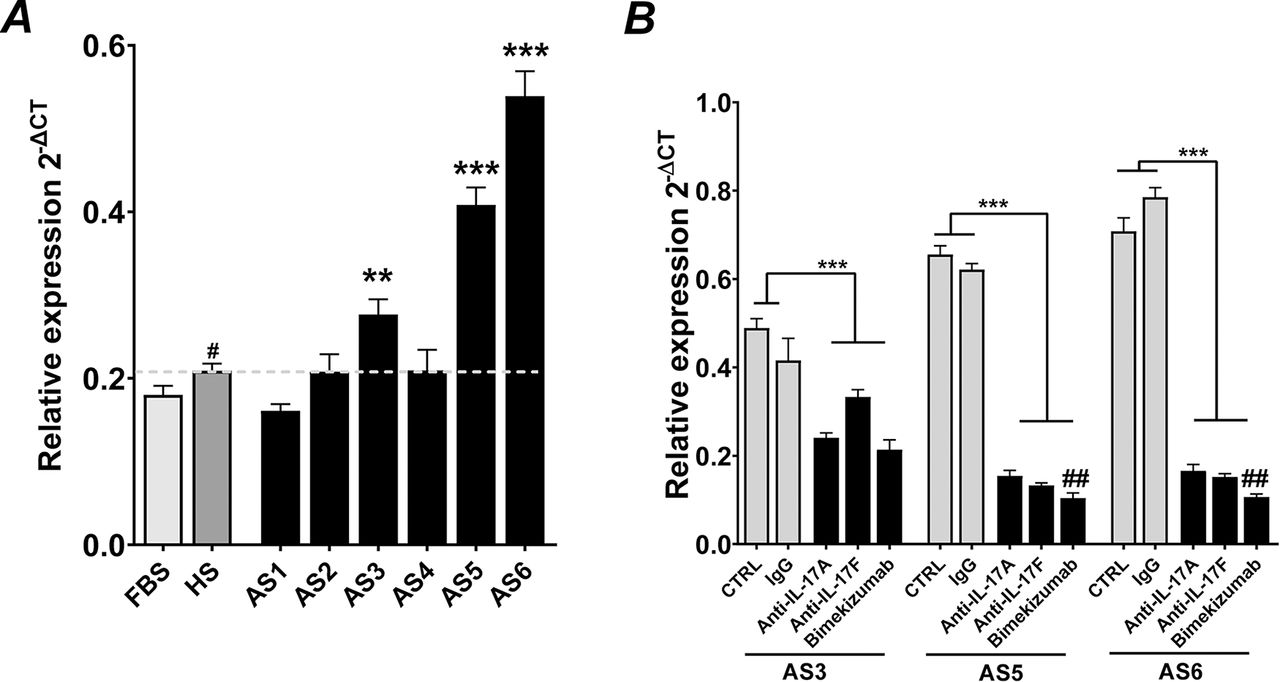
Osteocommitment of hPDCs by AS serum is suppressed by dual neutralisation of IL-17A and IL-17F
To mimic more closely the pathophysiology of the disease, we screened AS serum from six patients for their ability to induce expression of the osteocommitment marker RUNX2 (similar to that observed with T-cell supernatants). As seen in figure 3A, three cultures exhibited enhanced expression when compared to HS (p<0.001). To define the importance of IL-17A and IL-17F in the upregulation of RUNX2, sera were first treated with either control IgG antibody or antibodies specific to IL-17A, IL-17F or bimekizumab, prior to being added to hPDC cultures. Blocking IL-17A or IL-17F individually resulted in significant (p<0.01) decreases in RUNX2 expression (figure 3B); however, the effect was greatest with bimekizumab in two of three patients.
{kind=link}
{kind=link}
{kind=link}
Bimekizumab suppresses in vitro osteocommitment mediated by AS serum. (A) Expression of RUNX2 after stimulation of hPDCs with 10% AS patient-specific serum or 10% fetal bovine serum (FBS) or 10% HS (healthy human serum). Results are presented as the mean relative expression (2-ΔCT) ±SEM. ***p<0.001; **p<0.01; comparisons between serum from patient with AS and HS. ## p<0.01; # p<0.05; comparisons between FBS and HS by one-way ANOVA with Fisher’s LSD post hoc test (n=3). (B) Bimekizumab neutralisation of IL-17A and IL-17F in serum from patient with AS blocks osteocommitment as measured by RUNX2 expression over 48 hours. Results are the mean relative expression (2-ΔCT) ±SEM. *p<0.05; **p<0.01; ***p<0.001; (n=3). ANOVA, analysis of variance; AS, ankylosing spondylitis; hPDCs, human periosteum-derived cells; IL, interleukin; LSD, least significant difference.
DISCUSSION
We demonstrate herein for the first time that IL-17A and IL-17F potently enhance osteogenic differentiation and in vitro bone formation from human periosteal cells (hPDCs). Critically, these are the cells that are hypothesised by us, and others, to orchestrate pathological bone formation in AS. Blockade of both cytokines with bimekizumab in AS relevant assays using γδ or Th17 T-cell supernatants, or serum from patients with AS, affords deeper suppression of in vitro bone formation events than the blockade of either individually. This provides further scientific evidence for the dual blockade of IL-17A and IL-17F in patients with AS to prevent or suppress ankylosis of the spine.
Although we are the first to report on the pro-bone-forming capacity of IL-17A and IL-17F to human periosteal cells, others have investigated the role of IL-17A in periosteal biology using rodent models with conflicting results. Perhaps the most contradictory data come from a study conducted by Shaw et al,14 which reports the observation that IL-17A deficiency results in periosteal bone formation in response to K/BxN serum-transfer. Although an interesting observation, all mechanistic data presented were obtained through the analysis of calvarial osteoblasts in vitro, which are not equivalent to periosteal cells. Indeed, rat calvarial osteoblasts express primarily IL-17RB, IL-17RD and IL-17RE.15 Importantly, IL-17RD is reportedly able to negatively regulate IL-17A-induced activation of NF-κB,16 which may explain the divergent biology observed between hPDCs and rodent calvarial osteoblasts. Furthermore, Ono et al clearly demonstrate that while IL-17A stimulates osteoblastogenesis of precursor cells from long bone repair tissue, it does the opposite in calvarial osteoblasts.5 It is surprising that arthritis symptoms were not modulated in the study by Shaw et al,14 as it has previously been reported that genetic IL-17A deficiency17 or IL-17A antibody-mediated neutralisation18 suppresses arthritic disease in the same model. This aspect was not discussed by the authors but does warrant further investigation.
It has previously been reported that IL-17A can suppress the Wnt antagonist DKK1 in human mesenchymal stem cells.19 Mechanistically, Wnt signalling is known to be the master regulator of osteogenic differentiation. In agreement, IL-17 neutralisation herein resulted in enhanced DKK1 expression in hPDC cultures compared to those exposed to T-cell supernatants alone. Interestingly, recent clinical data detailing the treatment of PsA patients with secukinumab reported a progressive increase in serum DKK1 levels after 6 months of therapy.20 This is relevant to pathological bone formation, as AS serum DKK1 levels correlate with syndesmophyte formation.21 Furthermore, it has previously been shown that DKK1 expression is induced in response to TNF,22 which may be of relevance when interpreting data associated with anti-TNF therapy. Relevant to this, experiments conducted to deduce the composition of the biomimetic osteogenic conditions used herein describe TNF as having a negative effect on osteoblast gene expression.9 However, whether this effect is through the modulation of DKK1 expression is currently unknown.
To further elucidate the relevance of our findings to human AS, we screened serum from patients with AS for their capacity to induce osteocommitment of hPDCs. We subsequently found that serum IL-17A and IL-17F are capable of driving osteocommitment of human periosteal cells in vitro. Although not tested, this is likely to correlate to an enhanced capacity for matrix mineralisation and bone formation. To our knowledge, this is the first report that circulating cytokines are active and can drive disease biology. Interestingly, Uluçkan et al 23 have reported that circulating IL-17A suppresses intramedullary bone formation in a transgenic murine model of psoriasis. However, the authors did not report on periosteal bone formation or alterations in periosteal biology. It is also important to note that IL-17 cytokine action appears to be species dependent. Indeed, while IL-17F does not contribute significantly to inflammation in mice,24 we have recently demonstrated that it has qualitatively similar although quantitatively smaller pro-inflammatory effects in human in vitro models.10 This also limits the usefulness of mouse models in the study of IL-17F biology.
Finally, the data presented herein show IL-17F to be approximately equipotent to IL-17A in its ability to promote hPDC osteogenic differentiation, which is divergent from the lesser role that it appears to have in the inflammatory process. This suggests that dual neutralisation of IL-17A and IL-17F may be required to optimally suppress pathological periosteal bone formation observed in SpA.
Key messages
IL-17 signalling is a key regulator of inflammation in ankylosing spondylitis—but the role of IL-17 cytokines in pathological bone formation is poorly understood.
This study shows for the first time a direct effect of IL-17A and IL-17F on progenitor cells from the periosteum.
T-cell supernatants potently stimulate in vitro bone formation, which is blocked by dual neutralisation of IL-17F and IL-17A.
Inhibition of IL-17A and IL-17F may be required to optimally suppress pathological periosteal bone formation.
Acknowledgments
This work was funded by UCB Pharma. The authors would like to thank colleagues of UCB for their help, support and useful discussions. We would also like to thank Prof. Frank Luyten (KU Leuven, Belgium) for providing the hPDCs and Phil Stanley (UCB Pharma, UK) for providing expert statistical input.
REFERENCES
Footnotes
Contributors MS, SR, RK and SS contributed to the conception and design of the study. MS and AM were involved in the acquisition of data. PG, GM and RK provided scientific/clinical interpretation and patient material/data acquisition. MS, AM, DB, SS and SR contributed to the analysis and interpretation of data. SR supervised the study. All authors contributed to drafting and/or revising the manuscript.
Funding This work was funded by UCB Pharma.
Competing interests MS, AM, SS and DB are employees of UCB Pharma. PG has received honoraria from Medacta International. RK has received honoraria from Kyowa Kirin, Amgen and Clementia Pharmaceuticals, and consulting fees from Kyowa Kirin and Clementia Pharmaceuticals. SR is a former employee of UCB Pharma and has received consulting fees from Angitia Biopharmaceuticals.
Patient consent for publication Not required.
Ethics approval This study was approved by the NHS Research Ethics Committee (research authority: Yorkshire and the Humber—Leeds West Research Ethics Committee 16/YH/0137).
Provenance and peer review Not commissioned; externally peer reviewed.
Data availability statement All data relevant to the study are included in the article or uploaded as supplementary information.
Supplemental material This content has been supplied by the author(s). It has not been vetted by BMJ Publishing Group Limited (BMJ) and may not have been peer-reviewed. Any opinions or recommendations discussed are solely those of the author(s) and are not endorsed by BMJ. BMJ disclaims all liability and responsibility arising from any reliance placed on the content. Where the content includes any translated material, BMJ does not warrant the accuracy and reliability of the translations (including but not limited to local regulations, clinical guidelines, terminology, drug names and drug dosages), and is not responsible for any error and/or omissions arising from translation and adaptation or otherwise.