Article Text

Abstract
Objectives To evaluate the interferon (IFN) biomarkers sialic acid binding Ig like lectin 1 (SIGLEC1, CD169) and IFN-γ-inducible protein-10 (IP-10) in patients with primary Sjögren's syndrome (pSS).
Methods 31 patients fulfilling the American-European criteria for pSS were included. Disease activity was obtained by EULAR Sjögren's syndrome disease activity index (ESSDAI). SIGLEC1 expression on monocytes was analysed using flow cytometry. IP-10 concentrations were determined using Bioplex human Cytokine 27-plex kit. Spearman rank test (SRT) was used for correlation analysis and Mann-Whitney U (MWU) to test for differences between glandular and extraglandular manifestations.
Results An activated IFN system was detected by an upregulation of SIGLEC1 expression in 64.5% and by elevated serum level of IP-10 in 78.9% of our patients with pSS. In a subsequent analysis SIGLEC1 expression was found to be upregulated more frequently in patients with extraglandular manifestations (16/16, 100%) compared to patients with exclusively glandular involvement (4/15, 27%). SIGLEC1 expression could significantly discriminate between these two disease subgroups (p=0.0001, MWU) with a positive predictive value (PPV) of 80% for extraglandular disease. Moreover, the expression correlated with disease activity (p=0.005, r=0.54, SRT). Serum IP-10 levels neither differed significantly between glandular and extraglandular disease nor correlated with ESSDAI.
Conclusions Our results indicate that increased SIGLEC1 expression characterises patients with systemic involvement and high disease activity. Therefore, SIGLEC1 determination might be of value for subset definition, risk stratification and differential therapeutic considerations in pSS.
- Sjøgren's Syndrome
- Inflammation
- Disease Activity
- Cytokines
This is an Open Access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/
Statistics from Altmetric.com
Key messages
What is already known about this subject?
Interferons (IFNs) are known to contribute to the pathogenesis of primary Sjögren's syndrome (pSS) and the level of IFN-activation correlates with the disease activity of patients with pSS.
SIGLEC1 was identified in cell-specific transcriptome analysis as an IFN-regulated cell surface protein and was subsequently validated by flow cytometry as biomarker of disease activity in SLE.
What does this study add?
An upregulation of SIGLEC1 expression on peripheral monocytes was mainly detected in a subgroup of patients, that is simultaneously at higher risk for morbidity and mortality.
How might this impact on clinical practice?
SIGLEC1 might be useful to guide clinical decision-making, risk stratification or as pharmacodynamic marker in IFN-inhibitory therapies.
Since no clinically validated IFN-biomarker is available, SIGLEC1, with its detection by flow cytometry, is a prominent candidate that could easily be detected and implemented in clinical routine.
Introduction
Primary Sjögren's syndrome (pSS) is a chronic multifactorial autoimmune disease characterised by lymphocytic infiltration of the lacrimal and salivary glands, ultimately leading to glandular atrophy.1 Consequently, patients suffer from dryness of the mouth and eyes. A distinct subgroup of patients, characterised by systemic involvement and extraglandular manifestations, are at risk for higher mortality and morbidity.1–4 Anti-Ro/SSA and anti-La/SSB are key autoantibodies for the diagnosis of pSS and could be directly involved in the pathogenesis.1 However, the exact pathogenic principles remain unknown.
Over the last decade, growing evidence for type I and type II interferons (IFNs) in the pathogenesis of pSS has emerged.5–11 Studies using gene expression profiling of minor salivary glands and peripheral blood found a substantial upregulation of IFN-inducible genes (IFIGs) and an elevated type I IFN system was detected in 55% of patients with pSS.6 ,8 Additionally, the coexpression of five IFIGs (IFN score), consisting of MxA, IFI44, IFI44L, IFIT3, and LY6E, correlated with pSS disease activity.8 In a subsequent comparative study, IFN scores as determined by PCR correlated with IFN-related biomarkers, such as SIGLEC1 (CD169), CD64 and MxA by flow cytometry and enzyme linked immunosorbent assay (EIA), and MxA levels in monocytes correlated with the disease activity measured by ESSDAI.9
This study focuses on SIGLEC1, also known as sialoadhesin and CD169, a monocyte/macrophage-restricted cell surface protein with a variety of functions in the immune system, such as cell-to-cell interactions, antigen presentation and pathogen capture.12 SIGLEC1 was identified in cell-specific transcriptome analysis of patients with pSS7 and systemic lupus erythematosus (SLE),13 where it was one of the most prominently expressed genes within the IFN signature.13 SIGLEC1 expression in human monocytes is predominantly induced by type I IFN, while IFN-γ is only a weak inductor.14 ,15 Whereas SIGLEC1 expression on monocytes correlated with the disease activity in patients with SLE and was proposed as potential pharmacodynamic biomarker,13 ,16 ,17 it did not correlate with the disease activity of 28 patients with pSS presenting with inactive to mildly active disease (ESSDAI ≤6).9 Since an upregulation of SIGLEC1 expression is more prevalent among moderate to high disease activity in SLE,16 we hypothesize that a more systemic disease activity in pSS might also be reflected by an upregulation of SIGLEC1 expression.
Elevated levels of Interferon-γ-inducible protein 10 (IP-10), which is also inducible by type I IFN,18 were detected in different autoimmune diseases,19 including correlation with lupus activity cross-sectionally and longitudinally.16 ,18 ,20 Furthermore, elevated levels of IP-10 were found in patients with pSS related to T-cell infiltration into salivary glands.21 ,22 However, IP-10 has not yet been studied as a biomarker of systemic disease activity in pSS.
Evidence for an elevated interferon system in the pathogenesis of pSS is growing reflected by the facts that inhibiting IFN pathways is considered as a therapeutic option.23 However, surrogate markers for an activated IFN system that can be routinely used in clinical practice or for monitoring therapeutic effects have not yet been defined in pSS. In SLE, IP-10 and SIGLEC1 are candidates for implementation as IFN biomarkers, since both might be superior in monitoring disease activity compared to the current standard16 ,18 ,20 and were used in response to IFN-directed therapies such as sifalimumab24 and anifrolumab.25 This study, addressed the value of these two biomarkers in pSS with regard to disease activity and risk stratification of pSS subsets.
Patients and methods
Study participants
Thirty-six subsequent inpatient or outpatient patients with pSS at the Department of Medicine/Rheumatology and Clinical Immunology/Charité Berlin fulfilling the American-European criteria26 were screened. Exclusion criteria were a prednisolone dose >10 mg/day, pregnancy (n=3) and infections (n=2). Finally, 31 patients with pSS were included in this study. We did not apply additional selection criteria and since this study focused on the differences between glandular and extraglandular manifestations we included the same number of patients in each subgroup. Demographic data of patients is summarized in table 1.
Characteristics of patients with pSS and healthy controls
Disease activity of 25 patients was assessed according to the recommendation of the EULAR Sjögren's syndrome disease activity index (ESSDAI) at the point of blood sampling.27 Of these patients a detailed description including age, medication and SIGLEC1 expression can be found in online supplementary table S1. ESSDAI information of six patients was not available. Patients with ongoing or past organ involvement fulfilling the criteria of at least low activity in any organ domain of ESSDAI28 were considered as patients with extraglandular manifestations (n=16), while patients without systemic involvement were considered as solely suffering from glandular disease (n=15). Biological activity, indicated by parameters such as complement consumption or hypergammaglobulinemia, and constitutional symptoms were not taken into account for subset defintions. No patient with pSS had current or previous lymphoma.
supplementary table
As controls, 13 healthy donors (HD) were enroled. Patients with SLE fulfilling the American College of Rheumatology criteria served as controls (n=31).29 Twenty-two of 31 patients with SLE were treated with prednisolone (mean dose 5 mg/d), nine patients with hydroxychloroquine (mean dosage 333.3 mg/d), 13 with azathioprine (mean dose 95.5 mg/d) and four patients were receiving mycophenolat mofetil (mean dose 1693 mg/d). Disease activity scores of patients with lupus were not assessed. Written informed consent was obtained from all participants. The local ethics’ committee of the Charité Universitätsmedizin Berlin approved the study.
Detection of SIGLEC1 on monocytes using flow cytometry
Whole blood samples (0.2 mL; ethylene diamine tetra acetic acid (EDTA)) of patients with pSS, SLE and HD were immunophenotyped by flow cytometry. Erythrocyte lyses was performed using Becton Dickinson (BD) Pharm Lyse Solution (BD, Heidelberg, Germany) for 15 min at room temperature. After erythrolysis, cells were washed twice using MACS buffer (phosphate-buffered saline (PBS), 0.5% bovine serum albumin (BSA), 2 mM EDTA) (Miltenyi Biotec, Bergisch Gladbach, Germany). Prior to staining, the Fc receptor was blocked for 5 min at 4°C using FcR Blocking Reagent (Miltenyi Biotec). Cells were stained for 15 min at 4°C using 1:25 anti-CD14-APC-H7 (MOP9) (BD) for identification of monocytes and 1:50 anti-SIGLEC1-AF647 (7–239) (Biolegend, San Diego, USA) for the identification of SIGLEC1+ cells. The stained cells were washed using MACS buffer (PBS, 0.5% BSA, 2 mM EDTA) (Miltenyi Biotec). Shortly before measurement, 1 µL of 300 nM 4′,6-diamidino-2-phenylindole (Invitrogen, Carlsbad, USA) was added to exclude dead cells. The stained samples were analysed using a FACSCanto II (BD) flow cytometer within 1 hour. Standard operation procedure included daily use of Cytometer Setup and Tracking beads and Sphero Rainbow Calibration Particles-8 peaks beads (BD) in accordance with the quality control recommendation of the cytometer manufacturer. Signal readouts were the median fluorescence intensity (MFI) values for SIGLEC1 expression on CD14-positive monocytes, which was analysed with DIVA software V.6.0 (BD). A representative FACS plot is shown in figure 1.

Representative FACS plor of SIGLEC1 (CD169) expression on monocytes (CD14+) obtained from HD, pSS and SLE. The SIGLEC1 expression of patient with severe pSS disease acitivity is increased (ID19; ESSDAI 12; CD14/SIGLEC1+ 27.5%; median MFI 771), whereas the SIGLEC1 expression of patients with no pSS disease acitivity (ID6; ESSDAI 0; CD14/SIGLEC1+ 0.58%; median MFI 143) is comparable to HD (CD14/SIGLEC-1+ 1.02%; median MFI 144). Almost all monocytes of a patient with a SLE flare express SIGLEC1. ESSDAI, EULAR Sjögren's syndrome disease activity index; HD, healthy donor; MFI, median fluorescence intensity; pSS, primary Sjögren's syndrome; SIGLEC1, sialic acid binding Ig like lectin 1; SLE, systemic lupus erythematosus.
Detection of IP-10 in plasma samples of the donors using Bioplex
The levels of IP-10 were determined using the human Bioplex kit (Bio-Rad, Hercules, USA) with a sensitivity of 1.1 pg/mL. Frozen plasma samples of 19 patients with pSS, 26 patients with SLE and of 6 healthy donors were thawn and measured in accordance with the protocol recommended by the manufacturer and analysed using a Bio-Plex 200 system reader system (Bio-Rad, Hercules, USA).
Determination of threshold for SIGLEC1 and IP-10
The thresholds for SIGLEC1 and IP-10 were determined by receiver operating characteristics (ROC) curve analysis from HD versus patients with pSS. For SIGLEC1, a threshold of 260.5 MFI was determined resulting in a specificity of 100% with a sensitivity of 64.5%. For IP-10 we calculated a threshold of 175.8 pg/mL leading to a specificity of 100% with a sensitivity of 78.9%. A positive test was defined by a biomarker value above the calculated threshold with a specificity of 100%, indicating an activated IFN system.
Statistical analysis
Statistical analysis was performed using GraphPad Prism 6.0 (GraphPad, La Jolla, California, USA). Correlation between biomarkers and disease activity was calculated using Spearman rank test (SRT). Differences between patients with glandular and extraglandular manifestations were determined by Mann-Whitney U test (MWU). Differences in SIGLEC1 expression before and after the induction of hydroxychloroquine and prednisolne, respectively, were determined by Wilcoxon test for paired samples. p Values of less than 0.05 were considered significant.
Results
Patients with pSS show an enhanced IFN-activation compared to healthy controls
Initial analysis addressed whether the expression of SIGLEC1 on monocytes and IP-10 levels in plasma differ between HD and patients with pSS and SLE, respectively. Notably, both the levels of IP-10 (p=0.0005, MWU) and the expression of SIGLEC1 (p<0.0001, MWU) were significantly different between patients with pSS and HD. We found significantly higher levels of IP-10 in patients with SLE (p=0.0002), while we observed only a trend towards a higher SIGLEC1 expression in patients with SLE (p=0.052).
SIGLEC1, but not IP-10, can discriminate between glandular and extraglandular manifestations in pSS
Significant differences in SIGLEC1 expression on peripheral monocytes were observed between HD, patients with glandular (p=0.0176, MWU) and extraglandular manifestation (p<0.0001, MWU). Additionally, there was a significant difference of SIGLEC1 expression (p=0.0001, MWU) between both pSS subgroups (see figure 2A). In order to reduce confounding by medication, we performed a subgroup analysis with patients without medication and found a significant difference in SIGLEC1 expression (p=0.012, MWU) between patients with extraglandular and glandular disease (see figure 2B). Furthermore we investigated, if IFN inhibitory medications, such as prednisolone or hydroxychloroquine affect the SIGLEC1 expression.9 ,30 ,31 In a separate cohort we analysed the SIGLEC1 expression in patients with pSS and SLE before and after the introduction of prednisolone and hydroxychloroquine, respectively, and found a significant negative effect of both medication (see figure 3). Interestingly, the expression of SIGLEC1 was more reduced, in patients carrying higher baseline SIGLEC1 expression.

2/A Comparison of IP-10 and SIGLEC1 levels in healthy donors (HD), patients with glandular (gl.) and extraglandular (egl.) pSS and SLE 2/A. (MWU statistics was used to test for differences). 2/B. Subsequent analysis of pSS subgroups without medication (using MWU test). 2/C.Correlation analysis (using SRT) between disease activity (ESSDAI) of patients with pSS, SIGLEC1 expression (MFI) on monocytes and IP-10 in sera were performed. Red dots refer to patients with documented egl. manifestation, while black dots refer to patients suffering from gl. manifestation only. Dashed lines show calculated thresholds of SIGLEC1 and IP-10. 2/D. Comparison of SIGLEC1 expressions and levels of IP-10 in mild versus moderate to high disease activity. ESSDAI, EULAR Sjögren's syndrome disease activity index; MFI, median fluorescence intensity; MWU, Mann Whitney U Test; no med., no medication; ns, not significant; p values *, p<0.05; **, p<0.01; ***, p<0.001; ****, p<0.0001; r, correlation coefficient; SIGLEC1, sialic acid binding Ig like lectin 1; SRT, Spearman rank test.
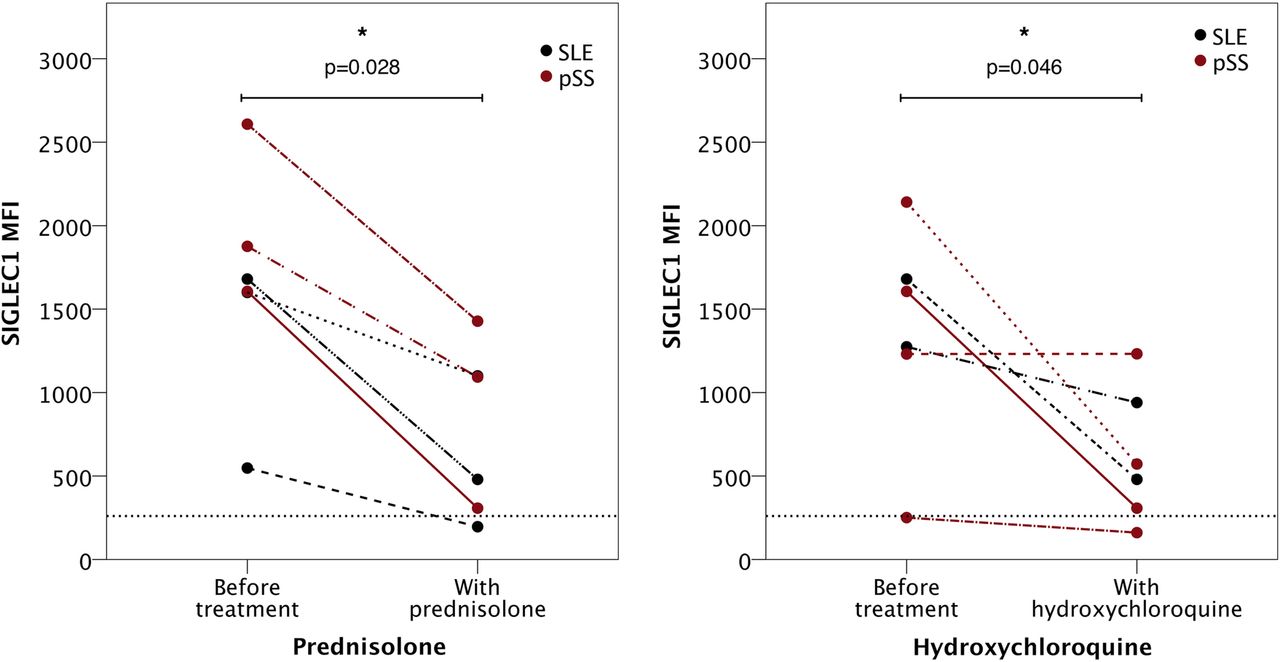
Mean fluorescence intensity (MFI) of SIGLEC1 expression before and after introduction of prednisolone or hydroxychloroquine. In a separate cohort, the MFI of SIGLEC1 on CD14+ monocytes of patients with either SLE (shown in black) or pSS (shown in red) was measured before and after the introduction of prednisolone or hydroxychloroquine. The average dose was 13.33 mg for prednisolone and 300 mg for hydroxychloroquine; the time period between measurements was ∼5.5 months for prednisolone and 5 months for hydroxychloroquine. Both treatments significantly reduced SIGLEC1 expression (p=0.028 and p=0.046, Wilcoxon test). The dashed line represents the calculated threshold of SIGLEC1 (260.5). pSS, primary Sjögren's syndrome; SIGLEC1, sialic acid binding Ig like lectin 1; SLE, systemic lupus erythematosus.
While levels of IP-10 did not differ between glandular and extraglandular manifestation (p=0.65, MWU), both pSS subgroups had significantly higher levels of IP-10 compared to healthy controls (gl. vs HD, p=0.001; egl. vs HD, p=0.007, MWU).
Subsequent analysis addressed if the degree of the IFN signatures are distinct in certain pSS subgroups based on the derived threshold. In the group with glandular manifestation we found that IP-10 was elevated in 80% (8/10) compared to SIGLEC1 in 26.7% (4/15). In the group of extraglandular manifestations IP-10 was elevated in 77.8% (7/9) compared to 100% (16/16) with an elevated SIGLEC1 expression. As a result, the positive predictive value (PPV) of an upregulated SIGLEC1 for extraglandular manifestations was 80%.
IFN activation and laboratory, clinical associations in pSS
Disease activity of our pSS cohort was assessed by ESSDAI providing global scores between 0 and 15 (see online supplementary table S1). Correlation analysis showed that SIGLEC1 correlated with ESSDAI (p=0.005, r=0.54) while IP-10 did not (p=0.58, SRT) (see figure 2C). In order to compare our results with the cohort of Maria et al, we analysed if the SIGLEC1 expression in patients with ESSDAI >5 differs from patients with ESSDAI <5 and found that ESSDAI >5 is associated with a significant upregulation of SIGLEC1 expression (p=0.0017, MWU) (see figure 2D).
Since IP-10 and SIGLEC1 are inducible by type I and II IFNs, we investigated whether SIGLEC1 expression and IP-10 levels interrelate, but we found no correlation (p=0.49, SRT). We further analysed, if patients lacked signatures of an activated IFN system but found only one patient who was neither positive for IP-10 nor for SIGLEC1 (see figure 4). Finally, we investigated if other laboratory values relate to ESSDAI domains. SIGLEC1 expression thereby correlated positively with constitutional symptoms of ESSDAI (p=0.004, r=0.55, SRT), whereas IP-10 did not show any association with clinical or laboratory parameters.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Distinct IFN pattern in pSS. Patients were ordered according their ESSDAI score. Yellow colour represents patients with an upregulation of SIGLEC1 and normal IP-10 levels. Blue colour indicates elevated levels of IP-10 but normal expression of SIGLEC1. Red colour indicates and upregulation of SIGLEC1 expression and elevated levels of IP-10. CNS, central nervous system; ESSDAI, EULAR Sjögren's syndrome disease activity index; IFN, interferon; PNS, periperhal nervous system; SIGLEC1, sialic acid binding Ig like lectin 1.
Discussion
Patients with pSS suffering from extraglandular manifestations are known to be at higher risk for all-cause mortality and certain of these manifestations contribute to serious morbidity.2–4 Therefore we analysed if an activated IFN system can identify those patients and found that all patients with extraglandular manifestations were characterised by upregulated SIGLEC1 expression. Its enhanced expression permitted a prediction of extraglandular manifestations in 80% (PPV) within the pSS cohort. This is the first study showing that an elevated IFN system detected by SIGLEC1 expression is prevalent in a pSS subgroup, that is simultaneously at higher mortality and morbidity risk. However, one patient with pSS (ID18, see online supplementary table S1) had only a mild upregulation of SIGLEC1 expression and showed a moderate disease activity with an ESSDAI of 13 (see figure 2C). This patient was taking 5 mg prednisolone daily, which is known to have an inhibiting effect on SIGLEC1 expression.13 ,32 The IFN-inhibitory effect of hydroxychloroquine is known for SLE and pSS.30 ,31 In this regard, we could confirm in a small longitudinal cohort that introduction of hydroxychloroquine and prednisolone reduces the IFN signature reflected by reduced SIGLEC1 expression. Thus, we assume that SIGLEC1 expression could be higher in this patient when assessed without the respective medication. Hydroxychloroquine was investigated in a randomised clinical trial (JOQUER trial) showing no efficacy in the treatment of inactive to mildly active patients with pSS (ESSDAI ≤6).33 In this context, it needs to be studied on a larger scale to which extent enhanced IFN activation can be modulated by hydroxychloroquine, as shown before and how this may support clinical decisions.
In the group of glandular manifestation, only four patients showed an upregulation of SIGLEC1. Two of them presented with moderate disease activity (with ESSDAI scores of 7 and 10, respectively), and three of them were positive for both autoantibodies (anti-Ro/SSA and anti-La/SSB). Noticeably, almost all other patients in the group of glandular manifestation presented with inactive disease and were only positive for anti-Ro/SSA antibody (see table 1). It has previously been described that anti-Ro/SSA und anti-La/SSB are both able to induce IFN production in pSS.34 Moreover, the simultaneous appearance of both autoantibodies (anti-Ro/SSA and anti-La/SSB) was associated with a lower rate of circulating plasmacytoid dendritic cells (pDC),35 which are considered as the main producers of IFN in pSS.7 ,35 These pDCs accumulate in the tissue in which they produce IFN.36 Hence, a higher frequency of migrated tissue pDC might lead to a higher activated IFN system, indicated here by an upregulation of SIGLEC1 expression and may explain a broader or more active organ involvement. Patients suffering from glandular disease may be at risk for further organ involvement when displaying an activated IFN system indicated by increased SIGLEC1 expression. Nevertheless, the current findings indicate that SIGLEC1 is a candidate for identifying patients with extraglandular manifestations.
In a previous study, the upregulation of type I IFN signature genes was positively correlated with the disease activity in pSS.8 ,9 MxA, but not SIGLEC1, could be shown to correlate with the disease activity in pSS.9 In contrast to Maria et al, we found a correlation of SIGLEC1 expression with disease activity. A possible explanation for this finding is, that low disease activity, with a low systemic IFN activation, might not be sufficient to upregulate SIGLEC1 expression. Patients of the cohort by Maria et al presented with low disease acitivity with the majority of patients with ESSDAI ≤6.9 Patients in our study presenting with moderate to high disease activity (defined by ESSDAI >5) showed a significant upregulation in SIGLEC1 expression and therefore might resemble a more systemic IFN activation. We conclude that our results are not contradictory to Maria et al, but rather extend that an activated IFN system is operative in pSS. Thus larger and longitudinal studies, including patients presenting with a wide range of disease activity, are needed to identify reliable IFN biomarkers for pSS.
In SLE, IP-10 was shown to correlate with disease activity cross-sectionally and longitudinally and therefore might be useful as an IFN biomarker in clincial practice in SLE.37 In pSS, IP-10 was shown to be involved in lymphocyte infiltration into glandular tissue of patients with pSS.21 Moreover, elevated levels of IP-10 in pSS were described recently by Howard Tripp et al.22 Thus, overlapping features in the pathogenesis of SLE and pSS and evidence of an involvement of IP-10 in the pathogenesis of pSS, led to the hypothesis that IP-10 might also serve as biomarker in pSS. This is the first study that investigated IP-10 as a biomarker of systemic disease avticity in pSS. Likewise to the study of Howard Tripp et al,22 we found that IP-10 was significantly upregulated in patients with pSS (80%) compared to HD. However, although SIGLEC1 and IP-10 are both induced by type I and II IFNs, unexpectedly, we could neither find a correlation of IP-10 to the disease activity nor observe a correlation between both markers.14 ,15 ,18 Gene expression data of in vitro stimulated human monocytes indicate quantitative and qualitative differences in the response signatures induced by IFN-α or with IFN-γ (see online supplementary figure S1).14 The kinetics are completely different for IP-10 and SIGLEC1 mRNA expression: while IP-10 is strongly upregulated very early after stimulation, the induction of SIGLEC1 is relatively delayed. Furthermore, there are indications that SIGLEC1 mRNA and protein expression is more stable compared to IP-10. The upregulation of IP-10 in response to IFN-α and IFN-γ is ∼50-fold higher compared to the upregulation of SIGLEC1. This could speak in favor that IP-10 is more acutely induced during the immune response and might be an explanation why there is a correlation to the flaring and waning disease course in SLE16 ,37 but not to the chronic disease course in pSS. Moreover, Smiljanovic et al14 and others showed that SIGLEC1 is mainly upregulated by IFN-α,32 while IP-10 equally responds to IFN-α and IFN-γ.14 Recently it was shown that type I and II IFN expression patterns may contribute to distinct clinical phe-notypes in pSS.10 Nezos et al investigated the IFN-signature in minor salivary glands (MSG) of patients with pSS which were complicated by lymphoma and found that IFNγ is more prevalent in MSG of patients with lymphoma, compared to patients without lymphoma. They proposed to use a ratio of IFN-α/IFN-γ in the diagnosis of salivary glands biopsies as a histopathological biomarker.10 Since IP-10 is regulated by IFN-γ, it would be interesting to study IP-10 in a pSS subset complicated by lymphoma.
supplementary figure
The current study has some limitations. First, we did not simultaneously analyse other biomarkers in pSS, such as MxA or CD64. In particular the difference between MxA and SIGLEC1 expression in the context of a higher disease activity would have been of interest. Second, the study did not enroll patients with pSS with central nervous or lymphoma manifestations which can contribute to an enhanced morbidity and mortality though both occur infrequently. There remains the possibility that type I IFN signatures may differ between additional subgroups of extraglandular disease which needs to be addressed by subsequent studies. Third, we cannot exclude that some patients taking hydroxychloroquine or prednisolone in the group of glandular disease would have an upregulation of SIGLEC1 expression without the respective medication. We compared the SIGLEC1 expression in a subsequent analysis of a smaller pSS subgroup without medication and found a significant difference in SIGLEC1 expression as well as a reduction of SIGLEC1 expression by prednisolone and hydroxychloroquine. It seems that SIGLEC1 expression is not only related to disease activity and with extraglandular disease but can also be modulated by therapeutic interventions. In addition to new insights into the immunopathogenesis, these aspects may have clinical implications including general and targeted treatment considerations, especially targeting IFN pathways.
References
Footnotes
TR and FS contributed equally.
Contributors TD proposed the study, contributed to the design of the study, clinical assessment, recruitment of patients; interpretation of the findings and drafting of the manuscript. TR contributed to clinical assessment and recruitment of patients, carried out the statistical analysis, the interpretation of the findings and preparation and drafting of the manuscript. FS performed the experimental analysis, contributed to the interpretation of the findings, to the manuscript preparation/drafting and interpretation of the findings. AL, KR and SJF contributed to the acquisition of data and contributed to the interpretation of the findings and to the manuscript preparation. GB, AR, FH, RB and AG contributed to interpretation of findings and critical review of the mansucript. All authors provided substantial contributions to the design or the acquisition, analysis, or interpretation of data of the study. All authors participated in revising the mansucript critically and approved the version to be published.
Funding Deutsche Forschungsgemeinschaft -DFG -SF650, IMMUNOBONE Do491/8-2, Do491/7-2,3.
Competing interests None declared.
Ethics approval Ethics committee of the Charité Berlin approved the study.
Provenance and peer review Not commissioned; externally peer reviewed.
Data sharing statement No additional data are available.